 2022, Vol. 40
2022, Vol. 40Institute of Oceanology, Chinese Academy of Sciences
Article Information
- ZHAO Ting, CAO Huidi, JIA Yanfen, HAN Xiaotian, YAN Tian, YU Rencheng
- Information standardization for typical toxic and harmful algae in China's coastal waters—a case study of Karenia mikimotoi
- Journal of Oceanology and Limnology, 40(6): 2178-2188
- http://dx.doi.org/10.1007/s00343-022-2169-z
Article History
- Received Apr. 6, 2022
- accepted in principle Jun. 9, 2022
- accepted for publication Jul. 8, 2022




2 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266100, China;
3 Eco-Environmental Monitoring and Research Center, Pearl River Valley and South China Sea Ecology and Environment Administration, Ministry of Ecology and Environment, PRC, Guangzhou 510610, China;
4 Qingdao Municipal Bureau of Ecology and Environment, Qingdao 266003, China;
5 Key Laboratory of Marine Ecology&Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
Harmful algal blooms not only harm marine life but also pose a threat to higher trophic organisms (including humans) through the food chain. At the same time, they also limit the development of the marine economy and are an important stress factor for the health of marine ecosystems and the safety of fishery resources.
Regarding current red tide events caused by toxic and harmful algae, many scholars have performed detailed studies (Zhou et al., 2001). For example, Gillibrand et al. (2016) conducted a systematic study on factors including the environment and vertical movement and migration of Karenia mikimotoi in the waters off the coast of Scotland from July to September 2006. Aoki et al. (2017) also investigated the causes of the K. mikimotoi tide that occurred in Iwanli Bay, eastern Japan, in 2014. However, these studies were aimed at a specific species in a certain period of time or in a certain region, including the location, area and economic loss of red tides. Knowledge of the long-term evolution law of red tides caused by different species is limited compared with that of their temporal and spatial distribution characteristics. From the perspective of environmental protection management, there is an urgent need for studies on the biogeographic distributions of diversified toxic and harmful algae, which require the introduction of an information management system as a comprehensive carrier of biological information to be managed.
An information management system (IMS) is an effective information carrier. It is the product of accounting, statistics, organizational theory, mathematical modeling, and other disciplines. Information management systems have been applied to marine biological information (Yamamoto et al., 2012). For example, for NCBI, Algae-Base, Bigelow, and other websites, most content includes data such as biological information and molecular information for algal species, but little information about the spatial and temporal distributions of alga-induced red tides is displayed, or it remains unavailable. The aims of this study are to digitize, unify, and standardize multidisciplinary information on different toxic and harmful algae and carry out standardized storage and management to rapidly disseminate the multidimensional information on toxic and harmful algae to researchers and decision managers.
In this paper, K. mikimotoi was selected as a typical toxic and harmful algal species. K. mikimotoi belongs to the order Dinoflagellata, widely distributed in temperate and tropical shallow waters, and is a typical ichthyotoxic red tide alga. In recent years, K. mikimotoi has caused several red tides in China's waters, the polluted area has gradually expanded, and the number of events has gradually increased. The toxins secreted by this microalga can seriously harm or kill fish and invertebrates, cause economic losses in fisheries, damage marine ecosystems, and even threaten human health through the food chain. For this paper, K. mikimotoi was selected as the research object, and its collected information was standardized to provide a reference for the collection and collation of biological information for other toxic and harmful algae in China's offshore waters.
2 MATERIAL AND METHOD 2.1 Information collectionBefore information standardization, information collection should be carried out. The purpose of information collection is to sort and classify available information, but it is also crucial to the construction of subsequent biological information management systems. In general, at the beginning of information collection, toxic and harmful algae distributed in China's offshore waters were determined as the target subjects. After establishing the research target subjects, the next step is to determine the detailed specific information needed about these toxic and harmful algal species. The basic process of information collection is shown in Fig. 1.

|
| Fig.1 Basic ideas for collecting information |
The basic biological information of K. mikimotoi, such as taxonomic status, toxins, morphological identification, pigment composition, lipid composition, and molecular characteristic sequences, were summarized and sorted. The biogeographic distribution information on offshore red tides of K. mikimotoi in China was sorted, the characteristics of red tide occurrence were analyzed, and standardization was completed. The obtained information, pictures, and other data were analyzed on the basis of their information characteristics and stored according to different data attributes to provide a basis for further database design.
2.2 Information acquisition 2.2.1 Document acquisitionFrom the perspective of basic biological information and biogeographic distribution information, the data items to be entered into the database during initial development (such as algal species name, density, life history, toxin, collection and culture, molecular biology information) are selected and collected, the standardization of data is performed, and the information standardization of the data required by the system is described.
2.2.2 Experimental acquisition 2.2.2.1 Microalgal cultureThe K. mikimotoi samples used in this work were provided by the Key Laboratory of Marine Ecology and Environmental Science, Institute of Oceanology, Chinese Academy of Sciences. All glassware was immersed in diluted HCl (10%) and washed several times with deionized water. Natural seawater from the East China Sea was filtered through a 0.45- m mixed-fiber membrane. An algal inoculum of 400 mL was transferred into culture vessels (5-L conical flasks containing 2 L of culture medium), the cultures were acclimatized with the medium of f/2 (Guillard and Ryther, 1962). The cultures were then acclimatized under an average irradiance of 150 μmol photons/(m2·s) with a light: dark regime of 12 h: 12 h with three replicates.
2.2.2.2 Morphological observationOptical microscope observations: K. mikimotoi was fixed with Lugo reagent, and an appropriate amount of algal liquid was taken for observation under an optical microscope (Nikon 1000).
Scanning electron microscope observation: a scanning electron microscope (SEM, TESCAN VEGA 3) was used to study the micromorphology of K. mikimotoi.
2.2.2.3 Pigment extraction and analysisHigh-performance liquid chromatography (HPLC, Waters e2695) was used for pigment composition analysis (Lai et al., 2012). K. mikimotoi were filtered through a GF/F filter membrane with a diameter of 47 mm. The obtained sample membrane (parallel samples P1 and P2) was wrapped with aluminum foil and stored in an ultra-low temperature refrigerator until analysis. The filter membranes were taken out from the low-temperature refrigerator and cut them into small pieces. Then transfer them into a brown sample bottle with a volume of 1.5 mL, and add 1 350-μL HPLC grade methanol and 150-μL internal standard mother liquor (8′-apo-beta-carotenal). Squeeze and crush the filter membranes to completely immerse them in the added methanol extract. Place the brown sample bottle in an ice water bath, after ultrasonic treatment for 5 min, suck the supernatant and filter through 0.2-μmol/L pore diameter needle type organic phase filter head, finally the filtrate was injected into a clean 1.5-mL sample bottle. After these steps, accurately suck 700-μL filtrate to another sample bottle, 140-μL ultrapure water was immediately add for analysis. The analysis method followed that of Zapata et al. (2000).
2.2.2.4 Lipid extraction and analysisThe experimental algal species were filtered by GF/F glass fiber membrane with a diameter of 47 mm. After filtration, the filter membranes were cut into small pieces and then 2.4 mL of solution (methyl tert-butyl ether (MTBE): MeOH=5:1) and a steel ball were added, the mixture was ground at 35 Hz for 4 min, and the process was repeated 4 times in an ice water bath for 5 min. The sample was centrifuged at 4 ℃ and 10 000 r/min for 15 min, and 1.75 mL of the supernatant was placed into an eppendorf (EP) tube for vacuum drying. Next, 200 μL of solution (dichloromethane (DCM): MeOH=1:1) was mixed, vortexed for 30 s, and ultrasonicated in an ice water bath for 10 min. Then, the sample was centrifuged at 4 ℃ and 13 000 r/min for 15 min, and 75 μL of supernatant was collected in an injection bottle.
For the detection method, refer to Dunn et al. (2011) and Want et al. (2010). Lipid components were detected using a SCIEX ExionLC™ AC system coupled to the SCIEX X500R Q-TOF-MS (AB SCIEX, USA). The chromatographic separation was conducted on a Phenomen Kinetex C18 (2.1 mm×100 mm, 1.7 μm). The mobile phase A was 40% Milli-Q water and 60% acetonitrile solution, containing 10-mmol/L ammonium formate, and the mobile phase B was 10% acetonitrile and 90% isopropanol solution. 50 mL of 10-mmol/L ammonium formate aqueous solution was added to every 1-L phase B. The separation was achieved with the following gradient elution condition: 40% B (0–12 min), linear increase to 100% B (12–13.5 min), 100% B (13.5–13.7 min), and finally decrease to 40% B (13.7–18 min). The injection volume was 1 μL (positive ion) and 2 μL (anion) and the flow rate was 0.3 mL/min for both positive and negative ion modes. The column temperature was 45 ℃.
2.2.3 Data storage and classificationWhen sorting the obtained information, it was classified according to its collectors and stored centrally. First, an original folder named after the algal species was established. Then, two subfolders of basic biological information and biogeographic information were set up, and several specific information subfolders were created in the two folders to classify and summarize the collected information in the form of tables or pictures.
2.2.4 Analysis of information characteristicsTo keep the data types entered into the database consistent, the data characteristics of the collected information were analyzed, the type of data was determined, and the data format was standardized, providing a reference for defining the data attributes in the database later.
3 RESULT AND DISCUSSION 3.1 Basic biological information 3.1.1 Names of algal speciesIn the early stage, there were many different names for the typical species studied, resulting in many aliases (former names), such as Gymnodinium sp., while the more recognized name is "Karenia mikimotoi", in which "mikimotoi" comes from the name of a Japanese farm owner (Lü et al., 2019) who funded research on the red tide of K. mikimotoi in his farming area. All relevant names were sorted in the study.
3.1.2 Classification statusHansen et al. (2000) separated this species from the genus Gymnodinium according to relevant data and established the genus Karenia. Bergholtz et al. (2006) synthesized the morphological characteristics, ribosomal DNA sequence (LSU rDNA) and pigment composition of K. armiger, and found the rDNA sequences of K. armiger and the genus Takayama are highly homologous. The authors considered the two taxa to belong to the same species. It is suggested to combine the three into a new family, Kareniaceae. By browsing the AlgaeBase website, we can find that Karenia mikimotoi belongs to the Eukaryota empire, Chromista kingdom, Harosa (supergroup stramenopiles-alveolates-rhizaria (SAR)) subkingdom, Harvard Infrakingdom, Miozoa phylum, Myzozoa subphylum, Dinophyceae class, Gymnodiniales order, Kareniaceae family, and Karenia genus, which are divided in great detail. The obtained classification status information will be used to establish the toxic and harmful alga directory on the information platform.
3.1.3 Life historyKarenia mikimotoi can adapt to various light, temperature, salinity, and nutrient conditions (Li et al., 2019). It can assimilate different forms of phosphates flexibly, and could take nutrients for growth autotrophically (by photosynthesis) and heterotrophically (by phagotrophy) simultaneously. K. mikimotoi propagates asexually mainly by division. No temporary cysts were found through molecular and morphological observation (Hansen et al., 2000). In culture experiments, it can be observed that some strains can get small cells through cell division. So far, there is no sufficient evidence to prove that K. mikimotoi can reproduce sexually, and its main mode of reproduction is asexual reproduction through cell division (Lü et al., 2019).
3.1.4 Toxin informationChen et al. (2015) and Li et al. (2017) conducted systematic studies of several toxins in K. mikimotoi and selected the three most common toxins: hemolytic toxins, cytotoxins, and fish toxins. Among them, hemolytic toxins (HTs) are the main cause of fish death during the red tides of K. mikimotoi. These toxins will cause pathological phenomena such as adjacent gill leaf adhesion, gill blood vessel rupture, blood cell exudation, and so on (Liu et al., 2015). Cytotoxins mainly consist of two toxins, gymnocin A and gymnocin B, which have similar characteristics to hemolytic toxins, but the difference is that hemolytic toxins mostly act on the cell membrane, while cytotoxins inhibit cell growth (Guo, 2014). However, many studies have also shown that hemolytic toxins and cytotoxins are not the main causes of death (Chang and Gall, 2013; Lin et al., 2016; Li et al., 2017). More in-depth research is needed on the toxins and toxicity mechanism of K. mikimotoi.
3.1.5 Morphological identificationThe optical microscopy images show that the algal body of K. mikimotoi is single-celled and greenish brown. At rest, only the dorsoventral surface can be seen, which is slightly flat (Fig. 2).

|
| Fig.2 Optical microscope observation |
According to morphological identification via scanning electron microscopy, K. mikimotoi cells are nearly round and slightly flat in dorsal and ventral view. The side, top, and bottom surfaces are hemispherical to widely conical, the upper cone has a groove at the top that extends upward from the upper end of the longitudinal groove to the top and reaches the back, and the bottom end of the lower cone has a depression (Fig. 3). The results of this morphological identification are consistent with records in the literature. It should be noted that during scanning electron microscope photography, it was found that the algal body was folded, which may be related to the growth period of the algae, fixation of the algal body and other factors. Their length was about 20–30 μm, and their width was about 16–30 μm. Basic on the Technical Specification for Red Tide Monitoring (HY/T 069-2055, 2005 (China's National Standard)), when the density were higher than 106 cells/L, it can be judged as the harmful algal blooms.

|
| Fig.3 Morphological identification of K. mikimotoi with SEM a–b. ventral side of K. mikimotoi; c–d. side, top, and bottom of K. mikimotoi. Scale bars=10 μm. |
The pigment compositions of P1 and P2 of the two control samples were detected by HPLC (Fig. 4). The results show that the pigments detected by the two samples were basically the same. The marker pigment of K. mikimotoi is fucoxanthin (FUCO), which includes 19′-butyryl FUCO and 19′-hexoyloxyfuco (hex-FUCO) (Suzuki and Ishimaru, 1992). In addition, K. mikimotoi also contains pigments such as didinoxanthin and 4-keto 19′ hexyl oxyfucophyll. Gyroxanthin diester pigment was also found in sample P1, which is a characteristic pigment of K. mikimotoi. Li et al. (2009) used this pigment and gyroxanthin to characterize K. mikimotoi in the large-scale red tide of dinoflagellates in the East China Sea, but this characteristic pigment peak was not obvious or absent in the two samples, which may be due to the low content of gyroxanthin diester pigment in this strain of K. mikimotoi. However, no obvious chlorophyll peak was detected during the experiment. Sample processing may take a long time, resulting in the degradation of chlorophyll.

|
| Fig.4 Pigment composition analysis of K. mikimotoi a. sample P1; b. sample P2. 1: but-FUCO like-1; 2: but-FUCO like-2; 3: but-FUCO; 4: FUCO; 5: Hex-FUCO like; 6: 4k-hex-FUCO; 7: Hex-FUCO; 8: unknown xanthophylls; 9: diad; 10: unknown xanthophylls; 11: gyroxanthin-diester. The peaks in blue boxes belong to group 10. |
The detected lipids mainly included triglyceride (TAG), phosphatidylglyceride (PG), monogalactose diglyceride (MGDG), phosphatidylethanolamine (PE), diglyceride (DAG), glucuronic acid glycolipid (GlcADG), diglyceride (DGDG), etc. (shown in Table 1). As a typical red tide alga producing fish toxins, K. mikimotoi is a haemolytic, cytotoxic species, and can produce substances with strong hemolytic activity, which are mainly glycolipids and unsaturated fatty acids (Yasumoto et al., 1990). MGDG and DGDG were haemolytic (Parrish et al., 1998). In addition, relevant substances such as lysophosphatidylcholine (LPC), lysophosphatidylethanolamine (LPE), and ceramide (Cer) were also found in this study. Lysophospholipids are single-chain fatty acyl phospholipid derivatives produced by the breaking of ester bonds at position 1 or 2 of phospholipids under the action of natural degradation or phospholipase (Li et al., 2018). Their components include measured LPC, LPE, and other substances.
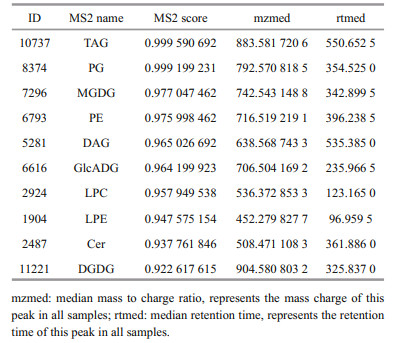
Molecular biology combined with morphological characteristics has gradually become a more common research method. Based on ultrastructural and molecular biological data, Hansen et al. (2000) distinguished the two species of K. mikimotoi from Gymnodinium aureolum and independently established a new genus for K. mikimotoi. Although the similarity of 18S rDNA sequences of different algal species in the same genus is very high, their sequences show great differences, especially in the ITS1 and ITS2 regions, which are more suitable for molecular identification among species. Kimura et al. (2015) constructed an RNA library by molecular means. Through analysis, it was found that some genes were similar to some toxin genes in other algal species, which provided a basis for studying the toxicological mechanism of K. mikimotoi. In addition, Zhang et al. (2009) also used loop-mediated isothermal amplification (LAMP) technology to identify the molecular sequence of K. mikimotoi from the South China Sea and achieved ideal results. According to this series of studies, information related to the identification of algal species at the molecular level has increasingly become an important tool and focus in algal research.
According to the relevant molecular sequences of algal species provided by the NCBI, the four regions of 18S rDNA, 28S rDNA, ITS1, and ITS2 and the sequences with obvious characteristics of the COX1 mitochondrial gene were selected (Table 2).

After collection of the required information, the information was integrated in the form of tables (Table 3). Other unused information was stored for later use.

According to the bulletin on the state of China's marine ecological environment, the bulletin on China's marine disasters, and a literature query, the distribution information of K. mikimotoi red tides in China's offshore areas is from south to north. From Zhejiang to Fujian waters are the mainly region of K. mikimotoi bloom (Liu et al., 2014; Ding and Zhang, 2018; Lü et al., 2019). Figure 5 shows many red tide events of K. mikimotoi in China's offshore areas. Red tides occurred almost every year from 1998 to the present (Fig. 5). Meanwhile, the reproduction of K. mikimotoi were distributed in the four sea areas of China, and the Donghai Sea has the most outbreaks (Fig. 6).

|
| Fig.5 The year and number of occurrences of K. mikimotoi along the coast of China |

|
| Fig.6 Karenia mikimotoi red tide distribution in various sea areas of China |
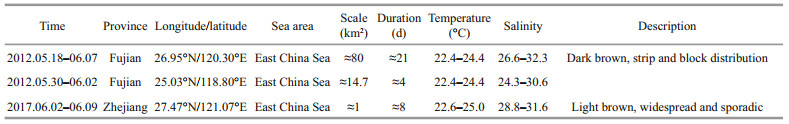
At the initial stage of the study, the names of typical toxic and harmful algae that induce red tides, the locations of red tides (longitude and latitude, province, sea area), scale, duration, a description of red tide characteristics, and the temperature and salinity of the sea area were sorted (Table 4). The reason for performing information collection is to determine the cause of red tides. Second, we sorted relevant information about the place of occurrence, including the sea area of occurrence and the longitude and latitude of the site. In addition, it was also necessary to record relevant information for each red tide, such as scale, duration, water temperature, and salinity of the sea area. Finally, it was also necessary to describe each red tide event, which will provide an intuitive representation for users.
Due to the wide range of information obtained, the information should be effectively classified and summarized during data storage. This paper classifies and arranges the collected information for K. mikimotoi into two parts: basic biological and biogeographic distribution information (Fig. 7).

|
| Fig.7 Information classification of K. mikimotoi |
For basic biological information, 18 field names, such as Chinese name, Latin name, classification status, size, morphological description, suitable growth temperature, pigment composition, and lipid composition information, were obtained according to the collected information (Table 5). Many characters were required for classification status, morphological description, pigment composition, lipid composition, molecular feature sequence information, and references. These fields were set to contain text for storage. Morphological images were stored in JPG format. Other information required fewer characters and was stored in character form.

For biogeographic distribution information, according to the collected information, the names of typical toxic and harmful algae inducing red tides, the occurrence locations of the red tides (longitude and latitude, province, and sea area), scale, duration, a description of red tide characteristics, and the temperature, salinity and references of the sea area were sorted, and the information characteristics were analyzed (Table 5). Except for the description of red tide characteristics and references, which required a certain amount of text, the data were stored as characters; other fields could be represented by a small number of characters or text, so they were set to the character type.
4 CONCLUSIONThis work collected and arranged the basic biological information of K.mikimotoi and obtained the data through auxiliary means (i.e., experiments). The biogeographic distribution information was collected comprehensively as a goal of this study. The harmful algal distribution map obtained through the secondary analysis of data will also be used as a reference for designing a biogeographic distribution function on the information platform in the future. According to basic biological information and biogeographic distribution information, the relevant information on the typical toxic and harmful alga K. mikimotoi was sorted, and information standardization was provided a template and method reference for the study of other toxic and harmful algae.
5 DATA AVAILABILITY STATEMENTThe datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Aoki K, Kameda T, Yamatogi T, et al. 2017. Spatio-temporal variations in bloom of the red-tide dinoflagellate Karenia mikimotoi in Imari Bay, Japan, in 2014: factors controlling horizontal and vertical distribution. Marine Pollution Bulletin, 124(1): 130-138.
DOI:10.1016/j.marpolbul.2017.07.019 |
Bergholtz T, Daugbjerg N, Moestrup Ø, et al. 2006. On the identity of Karlodinium veneficum and description of Karlodinium armiger sp. Nov. (Dinophyceae), based on light and electron microscopy, nuclear-encoded LSU rDNA, and pigment composition. Journal of Phycology, 42(1): 170-193.
DOI:10.1111/j.1529-8817.2006.00172.x |
Chang F H, Gall M. 2013. Pigment compositions and toxic effects of three harmful Karenia species, Karenia concordia, Karenia brevisulcata and Karenia mikimotoi (Gymnodiniales, Dinophyceae), on rotifers and brine shrimps. Harmful Algae, 27: 113-120.
DOI:10.1016/j.hal.2013.05.005 |
Chen B H, Xie E Y, Gao Y H, et al. 2015. Toxic effects of red tide caused by Karenia mikimotoi on marine organisms. Journal of Fujian Fisheries, 37(3): 241-249.
(in Chinese with English abstract) DOI:10.14012/j.cnki.fjsc.2015.03.010 |
Ding G M, Zhang S F. 2018. Ecological characteristics and the causes of Karenia mikimotoi bloom in the Sansha Bay in 2012. Haiyang Xuebao, 40(6): 104-112.
(in Chinese with English abstract) DOI:10.3969/j.issn.0253-4193.2018.06.010 |
Dunn W B, Broadhurst D, Begley P, et al. 2011. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nature Protocols, 6(7): 1060-1083.
DOI:10.1038/nprot.2011.335 |
Gillibrand P A, Siemering B, Miller P I, et al. 2016. Individual-based modelling of the development and transport of a Karenia mikimotoi bloom on the North-west European continental shelf. Harmful Algae, 53: 118-134.
DOI:10.1016/j.hal.2015.11.011 |
Guillard R R L, Ryther J H. 1962. Studies of marine planktonic diatoms: I. Cyclotella nana hustedt, and Detonula confervacea (cleve) gran. Canadian Journal of Microbiology, 8(2): 229-239.
DOI:10.1139/m62-029 |
Guo H G. 2014. Study of Purification and Characterization of Hemolytic Toxin and Cytotoxin from Karenia mikimotoi. Xiamen University, Xiamen. (in Chinese with English abstract)
|
Hansen G, Daugbjerg N, Henriksen P. 2000. Comparative study of Gymnodinium mikimotoi and Gymnodinium aureolum, comb. nov. (=Gyrodinium aureolum) based on morphology, pigment composition, and molecular data. Journal of Phycology, 36(2): 394-410.
DOI:10.1046/j.1529-8817.2000.99172.x |
HY/T 069-2005. 2005. Technical specification for red tide monitoring. State Oceanic Administration.
|
Kimura K, Okuda S, Nakayama K, et al. 2015. RNA sequencing revealed numerous polyketide synthase genes in the harmful dinoflagellate Karenia mikimotoi. PLoS One, 10(11): e0142731.
DOI:10.1371/journal.pone.0142731 |
Lai J X, Yu Z M, Song X X, et al. 2012. Pigment-based assessment of the response of phytoplankton to nutrient addition at the Changjiang Estuary. Marine Sciences, 36(5): 42-52.
(in Chinese with English abstract) |
Li J, Glibert P M, Zhou M J, et al. 2009. Relationships between nitrogen and phosphorus forms and ratios and the development of dinoflagellate blooms in the East China Sea. Marine Ecology Progress Series, 383: 11-26.
DOI:10.3354/meps07975 |
Li X D, Yan T, Lin J N, et al. 2017. Detrimental impacts of the dinoflagellate Karenia mikimotoi in Fujian coastal waters on typical marine organisms. Harmful Algae, 61: 1-12.
DOI:10.1016/j.hal.2016.11.011 |
Li X D, Yan T, Yu R C, et al. 2019. A review of karenia mikimotoi: bloom events, physiology, toxicity and toxic mechanism. Harmful Algae, 90: 101702.
DOI:10.1016/j.hal.2019.101702 |
Li Z, Gu K R, Zhang H C, et al. 2018. Progress in preparation, separation, determination and application of lysophospholipids. China Oils and Fats, 43(6): 132-136, 143.
(in Chinese with English abstract) DOI:10.3969/j.issn.1003-7969.2018.06.028 |
Lin J N, Yan T, Zhang Q C, et al. 2016. Effects of Karenia mikimotoi blooms on antioxidant enzymes in gastropod abalone, Haliotis discus hannai. Marine Sciences, 40(6): 17-22.
(in Chinese with English abstract) DOI:10.11759/hykx20150330001 |
Liu G Y, Ge K, Song L, et al. 2015. The research trend of Karenia mikimotoi. Marine Sciences, 39(9): 117-122.
(in Chinese with English abstract) DOI:10.11759/hykx20130702002 |
Liu Z G, Wang J H, Cai G, et al. 2014. Distribution of Karenia mikimotoi and rules of its causing red tide. Territory and Natural Resources Study, (1): 38-41.
(in Chinese with English abstract) DOI:10.3969/j.issn.1003-7853.2014.01.015 |
Lü S H, Cen J Y, Wang J Y, et al. 2019. The research status quo, hazard, and ecological mechanisms of Karenia mikimotoi red tide in coastal waters of China. Oceanologia et Limnologia Sinica, 50(3): 487-494.
(in Chinese with English abstract) DOI:10.11693/hyhz20181000255 |
Parrish C C, Bodennec G, Gentien P. 1998. Haemolytic glycoglycerolipids from Gymnodinium species. Phytochemistry, 47(5): 783-787.
DOI:10.1016/S0031-9422(97)00661-4 |
Suzuki R, Ishimaru T. 1992. Characteristics of photosynthetic pigment composition of Gymnodinium mikimotoi MIYAKE et KOMINAMI EXODA. Journal of Oceanography, 48(4): 367-375.
DOI:10.1007/BF02234015 |
Want E J, Wilson I D, Gika H, et al. 2010. Global metabolic profiling procedures for urine using UPLC-MS. Nature Protocols, 5(6): 1005-1018.
DOI:10.1038/nprot.2010.50 |
Yamamoto H, Tanaka K, Fujikura K et al. 2012. BismaL: biological information system for marine life and role for biodiversity research. In: Nakano S I, Yahara T, Nakashizuka T eds. The Biodiversity Observation Network in the Asia-Pacific Region. Springer, Tokyo. p. 247-256, https://doi.org/10.1007/978-4-431-54032-8_18.
|
Yasumoto T, Underdal B, Aune T, et al. 1990. Screening for hemolytic and ichthyotoxic components of Chrysochromulina polylepis and Gyrodinium aureolum from Norwegian coastal waters. Toxic Marine Phytoplankton, 45: 436-440.
|
Zapata M, Rodríguez F, Garrido J L. 2000. Separation of chlorophylls and carotenoids from marine phytoplankton: a new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Marine Ecology Progress Series, 195: 29-45.
DOI:10.3354/meps195029 |
Zhang F Y, Xu Z L, Ma L B, et al. 2009. Loop-mediated isothermal amplification (LAMP) establishment for detection of Karenia mikimotoi (Dinophyceae). Acta Oceanologica Sinica, 31(6): 170-175.
(in Chinese with English abstract) DOI:10.3321/j.issn:0253-4193.2009.06.019 |
Zhou M J, Zhu M Y, Zhang J. 2001. Status of harmful algal blooms and related research activities in China. Chinese Bulletin of Life Sciences, 13(2): 54-59, 53.
(in Chinese with English abstract) DOI:10.3969/j.issn.1004-0374.2001.02.002 |