 2022, Vol. 40
2022, Vol. 40Institute of Oceanology, Chinese Academy of Sciences
Article Information
- FANG Guangjie, YU Haolin, WANG Xinmeng, SHENG Huaxiang, TANG Yanli, LIU Changdong, CHEN Chuanxi, LIANG Zhenlin
- Impact of summer moratorium on set-net fishery in Haizhou Bay, China
- Journal of Oceanology and Limnology, 40(4): 1678-1691
- http://dx.doi.org/10.1007/s00343-021-0479-1
Article History
- Received Dec. 22, 2020
- accepted in principle Mar. 30, 2021
- accepted for publication Sep. 6, 2021




2 Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China;
3 Marine College, Shandong University, Weihai 264200, China
Fishery stocks are declining in different degrees worldwide because of overexploitation and other human-induced environmental changes (Jackson et al., 2001; Davies and Baum, 2012). The risks of the fishery industry in China are highly urgent (Shen and Heino, 2014). Haizhou Bay is located in the southern Yellow Sea near the province of Shandong, China, with an area of about 820 km2. The shape of the bay likes a trumpet; many rivers along the coast input a great quantity of nutrients into the sea, which creates an excellent feeding and spawning ground for fishery species (Pang et al., 2015). As one of China's eight principal fishing grounds, fishery resources (e.g., small yellow croaker Larimichthys polyactis, hairtail Trichiurus lepturus, and mackerel Scomberomorus niphonius) in Haizhou Bay is very rich before the 1990s (Su et al., 2015). However, due to overfishing and habitat destruction, the main economic species of set-net fishery in Haizhou Bay are in a state of overexploitation (Su et al., 2015).
To protect fishery resources by ensuring their reproduction in the spawning season, in 1995, China implemented a seasonal closure policy called "summer moratorium of marine fishing" (SMMF). SMMF is a "command and control" policy, which is a command issued by the government, and set standards and control performance by monitoring and requiring adherence to that standards. Because of significant differences in sea regions and fishing gears, the implementation of SMMF differs correspondingly. In the past twenty years, the policies of SMMF adjusted considerably from two months to four months, and now, SMMF is carried out from May 1st to September 1st for the most fishing gears in Haizhou Bay. At present, SMMF has been one of China's most effective fishery management policies because of essential goals and strict implementation (Yue et al., 2016; Yan and Lu, 2020). Many studies showed that some target stocks' catch rates and fish sizes have increased with different degrees (Yan et al., 2006; Yu et al., 2017). In the past twenty years, the impact of SMMF was studied by researchers mainly focused on the single-species fisheries (Zou et al., 2016), the change of community structure in one or two years (Yan et al., 2016; Yu et al., 2018). However, few studies focusing on the impact of SMMF on the small-scale fishery in China and provide suggestions for policy improvements.
Millions of people in developing countries depend on small-scale fisheries, such as Chinese artisanal fisheries, for their food and income (Sowman, 2006; Yang et al., 2011). The set-net fishery is an important part of small-scale fisheries in China, which relies on water flow to catch fish, belonging to "low impact and fuel efficient" fishing gear (Suuronen et al., 2012). Due to its advantages of low energy consumption, low technical requirements (low demand for high-tech equipment), low cost, and stable output; the set-net fishery has developed rapidly. According to statistics, their fishing production ranks 3rd in the total fishing production in the Yellow Sea, China, which is a crucial fishing gear for subsistence fishers (Sun et al., 2012).
The set-net fishery is a multi-species fishery; it is helpful to evaluate the changes of targeted species to analyze the changes of fishery community compositions. Six principal species, pinkgray goby (Amblychaeturichthys hexanema), small yellow croaker (Larimichthys polyactis), white-hair rough shrimp (Trachypenaeus curvirostris), swimming crab (Portunus trituberculatus), mantis shrimp (Oratosquilla oratoria), and short arm octopus (Octopus ocellatus) were identified based on catch data sets. A. hexanema is one of the most abundant fish with no economic value in the set-net fishery due to rapid growth and early sexual maturity (Wu and Zhong, 2008). Its spawning time is from April to June. The other five species are the main target species because of their high economic values and yields. L. polyactis grows faster during summer and its main hatching period is from May to August (Pei et al., 2021). T. curvirostris is an important commercial fishing species in Northeast Asia, and its main spawning period is from July to August (Chen, 1991). P. trituberculatus is a sizeable economic crab in the coastal waters of China; its hatching period is mainly from June to July (Xu et al., 2021). O. oratoria is a perennial crustacean distributed widely on the coast of China, and its spawning time is mainly from May to July (Geng et al., 2019). O. ocellatus is the most yield Cephalopoda in the Yellow Sea, and its hatching time is primarily from March to May (Jin et al., 2018).
In the present study, we estimated the dynamics of set-net fishery in Haizhou Bay, China with eight years for the first time, and investigated the impact of SMMF on the fishery communities. The primary objectives were (1) to analyze the changes of biological characteristics and community compositions in the set-net fishery from 2011 to 2018; (2) to summary the effects of SMMF on the principal species, groups and total communities, and provide suggestions for the subsequent policy improvement.
2 MATERIAL AND METHOD 2.1 Study area and samplingThe study was carried out along the Haizhou Bay, Yellow Sea, China from 2011 to 2018 (Fig. 1). Three sites (A, B, and C) were deployed in the traditional set-net fishing region, and the sites had similar fishing operations and environmental conditions (Tang et al., 2018). In these areas, the depth of seawater was about 20 m, and the seafloors are mud. The distances of the three stations to the fishing ports are about six nautical miles, and local fishers have similar working time and operating habits. The fishing nets were double-staked vertical bar set-nets (Fig. 1), which are widely used for small boats in Haizhou Bay. There was no change in fishing nets in eight years. The principal dimensions of the nets differed by stations slightly, which were 46.00 m × 38.40 m, 47.80 m × 38.24 m, and 56.40 m × 39.48 m (net mouth circumference × net length) for stations A, B, and C, repsectively. The material of nets was polyethylene, and the cod-end mesh size was 17 mm. There were generally 10 to 20 same nets fixed in the same station, and fishers collected catches from cod-ends every day. Every year, to detect the impact of SMMF on the set-net fishery, four times of sampling surveys before SMMF and after SMMF during two months were conducted, respectively. Meantime, about 10 kg of catches were randomly sampled for every station in the fishing ports, and all samples were taken to the laboratory and sorted, identified, and enumerated, and the total length and weight was measured for all individuals captured.

|
| Fig.1 Map of the study area with three sites (A, B, and C) and the sketch of the set-net |
Two data sets were collected to analyze the change in catch per unit effort (CPUE): the set-net fishery's biological characteristics and community structure from 2011 to 2018. One data set is fishing logs recorded by fishers daily during the study periods, containing fishing dates, nets, and daily catch weights for each station. We designed the fishing logs and trained fishers to record data systematically. The second data set is the compositions of sampling catches captured from fishing ports, including the biological characteristics of all individuals.
2.2 Data analysisDaily fishing catch weights were standardized to remove the influence of different net mouth areas (Tang et al., 2014), and CPUE was estimated as the catch weight per standard net per day (Su et al., 2015; Tang et al., 2018). The computational formula for the standardized CPUE was as follows:
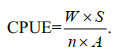 (1)
(1)In this equation, W is the daily catch weight (kg); S, the standard net mouth area (100 m2); n, the daily numbers of set-nets; and A, the actual net mouth area (m2). The unit for CPUE is kilogram per net per day (kg/(net·d)). Mann-Whitney test was used to compare the mean CPUE before and after SMMF in eight years.
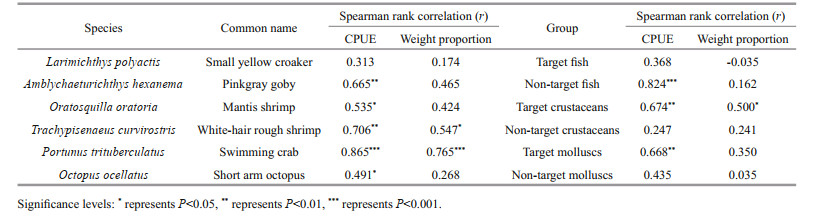
Spearman rank correlation was used to identify whether six species and six groups were responsible for the changes in the fishery community (Jiang et al., 2009). The CPUE, weight proportion of six species and six groups were obtained to analyze the correlations with the CPUE of the total catches from 2011 to 2018. Six principal species were selected to ascertain the temporal variations of fishery communities and the influence of SMMF on the set-net fishery because of their high abundances and economic values. The mean total length, CPUE and the mean weight proportion in the total catches of the six species were estimated. Meanwhile, the principal species and species groups were used to estimate the changes of communities and the effects of SMMF (McClanahan, 2014). Total species were divided into six groups: target fish group, non-target fish group, target crustaceans group, non-target crustaceans group, target molluscs group, and non-target molluscs group. Fishery-target species are targeted or highly valued by commercial fishing, and non-target species are all other species. These groupings aimed to determine whether target species exhibited different temporal trends in relative abundance compared to non-target species.
Most diversity indices are sensitive to specific aspects of structural or functional change in the community, and each has particular advantages and disadvantages (Rice, 2000). Three diversity indices were calculated to compare the changes of community compositions in study years. The Shannon-Wiener diversity index (H′) was used for species diversity (Ludwig and Reynolds, 1988):
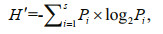 (2)
(2)where s is the number of the total species, and Pi is the proportion of the total sample represented by species i.
Simpson diversity index (D) reflected the dominance of dominant species (Simpson, 1949):
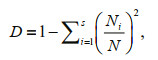 (3)
(3)where s is the number of the total species, and Ni is the proportion of total sample represented by species i.
Pielou's evenness index (Jʹ) was used for evaluating the distribution uniformity of different species in the community of the set-net fishery (Pielou, 1966).
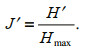 (4)
(4)In this formula, Hʹ is the Shannon-Wiener diversity index; Hmax=log2s mean the maximum diversity possible; and s is the number of total species.
We used non-metric multidimensional scaling (NMDS), which is a reduced-space ordination method based on a Bray-Curtis dissimilarity matrix, to analyze community composition before SMMF and after SMMF with software package PRIMER 6 (Clarke and Gorley, 2006). Analysis of similarity (ANOSIM) is a nonparametric method used to test whether the difference between groups is significantly greater than the difference within groups (Clarke, 1993). Permutation-based analysis of variance (PERMANOVA) is a nonparametric method proposed to test the difference between two or more groups of objects by decomposing the sum of the squares of their distances (Anderson, 2001). ANOSIM and PERMANOVA were calculated to test the differences among the communities before and after SMMF from 2011 to 2018. The average contribution of individual sampling species to the overall Bray-Curtis distances was estimated using the similarity percentage (SIMPER) analysis (Clarke and Warwick, 1994). SIMPER was used to compare the community dissimilarity between two periods (before and after SMMF) and analyze the similarity in the same period, and then the contributions of species for similarity and dissimilarity were calculated (Clarke, 1993). ANOSIM, PER-MANOVA, and SIMPER were performed in R 3.5.2 using the "vegan" package.
3 RESULT 3.1 Species compositionsA total of 141 species belonging to 67 families were identified from 192 fishing samples in eight years. Among these, 41 families of fish, 21 families of crustaceans, and 5 families of molluscs were observed. The whole species were divided into six groups based on their economic values (Table 1). The non-target fish group was the most abundant group for the mean weight proportion (46.6%) before and after SMMF. The mean weight proportions of the target fish group, non-target fish group, target crustaceans group, and non-target crustaceans group were over 10%.

The CPUE of the five principal species was positively correlated to the total CPUE in the set-net fishery from 2011 to 2018 (Table 2), which suggested that these species could represent the whole community compositions. Before SMMF, the mean total length of L. polyactis, T. curvirostris, P. trituberculatus, and O. ocellatus showed substantial decreasing trends (over 10%) in eight years (Fig. 2). After SMMF, the mean total length of P.trituberculatus and A. hexanema increased slightly, while the other four species had varying degrees of decline (Fig. 2).
|

|
| Fig.2 Changes of mean total lengths of six species: Larimichthys polyactis, Trachypenaeus curvirostris, Portunus trituberculatus, Amblychaeturichthys hexanema, Oratosquilla oratoria, and Octopus ocellatus during two periods (before and after summer moratorium of marine fishing) from 2011 to 2018 The dotted line was fitted using an ordinary least squares model. |
The mean weight proportion of the target and nontarget groups showed varied inter-annual fluctuations. Before SMMF, the developing trends of the weight proportion for the target and non-target groups were opposite (Fig. 3). For fish, the weight proportion of the non-target fish group was significantly higher than that of the target fish group in eight years. Although an over 10% decreasing trend for the mean weight proportion of the non-target fish group was observed, the weight proportion of the target fish group was stable. As for crustaceans, the weight proportion of the non-target group was higher than the target group. A significantly increasing trend of the weight proportion for the non-target group and a slightly decreasing trend for the target groups were found, respectively. About molluscs, the weight proportion of the target group showed a sharp decline, and the non-target group surpassed the target group after 2017.

|
| Fig.3 Weight proportion of six groups during two periods from 2011 to 2018 The dotted line was fitted using an ordinary least squares model. SMMF: summer moratorium of marine fishing. |
After SMMF, for fish, the mean weight proportion of the non-target group was nearly twice that of the target group in the study years, and slight decreases of the weight proportion for both the target and non-target groups were observed. As for crustaceans, the mean weight proportion of the target group increased by over 10%, and the weight proportion of the non-target group decreased. Compared with the tendencies of these two groups before SMMF, absolutely opposite directions were exhibited. About molluscs, the mean weight proportion of the target group decreased by more than 50%, and the decline also occurred in the non-target group.
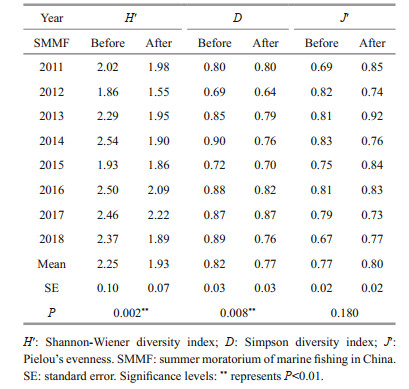
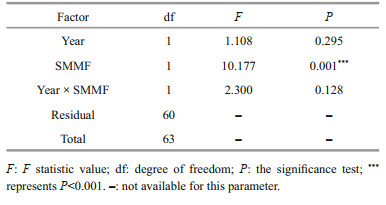
For three diversity indices, no significant temporal trend was found (Table 3). ANOSIM indicated that community compositions among eight years were similar (P > 0.05). PER-MANOVA showed that the factor "Year" did not influence the fishery communities significantly (Table 4). SIMPER revealed that the similarity of communities among years before SMMF was 77.54%, and the contributions of L. polyactis, A. hexanema, and P. trituberculatus were over 10%. The similarity after SMMF among years was 76.68%, and the contributions of A. hexanema and P. trituberculatus were over 10%.
|
|
For the six principal species, the mean CPUE after SMMF from 2011 to 2018 was higher than before SMMF except for O. ocellatus (Fig. 4). Significant differences were observed before and after SMMF for L. polyactis, T. curvirostris, P. trituberculatus, and O. ocellatus. Meanwhile, the mean weight proportion of the six principal species after SMMF was higher than before SMMF except for O. ocellatus (Fig. 4). For the six groups, the target fish and crustaceans group showed higher mean weight proportions after SMMF in eight years than before SMMF, while the two non-target groups were converse (Fig. 3). As for molluscs, the mean weight proportion of the two groups before and after SMMF were highly consistent (Fig. 3).

|
| Fig.4 Mean catch per unit efforts (CPUE, kg/(net·d)) and mean weight proportion of CPUE for six species: Larimichthys polyactis, Trachypenaeus curvirostris, Portunus trituberculatus, Amblychaeturichthys hexanema, Oratosquilla oratoria, and Octopus ocellatus before and after summer moratorium of marine fishing (SMMF) from 2011 to 2018 Asterisks represent significant differences in CPUE before and after SMMF at the following alpha levels: *: 0.05, **: 0.01. |
For diversity indices (H′, D, J′), the three indices before and after SMMF during study periods were shown in Table 3. Inter-annual variations for three diversity indices were not evident (P > 0.05), but significant differences of mean H′ and D before and after SMMF were found (P < 0.01). H′ and D were higher before SMMF than after SMMF, and J′ was higher after SMMF than before SMMF (Table 3). The NMDS analysis showed that the entire communities before and after SMMF were two distinct groups (Fig. 5). ANOSIM indicated that community compositions between the two periods were significantly different (P < 0.01). PER-MANOVA also showed that SMMF was a significant factor (P < 0.001) that influence the communities of the set-net fishery (Table 4). SIMPER analysis revealed that dissimilarity between the two periods was 38.76%, and the contribution of L. polyactis was over 10%.

|
| Fig.5 Non-metric multidimensional scaling diagram showing community structure differences between two periods (before and after the summer moratorium of marine fishing) from 2011 to 2018 |
In our eight study periods, losses of the mean total length for the five principal economic species were observed, especially for L. polyactis (Fig. 2). Many reasons could explain these phenomena. For one thing, overfishing may induce the rapid evolution of marine organisms (e.g., individual miniaturization) and hinder population recovery (Kuparinen and Merilä, 2007). L. polyactis, known as one of China's four major marine economic fishes in the 20th century, is an important fishing species. However, under the high fishing pressure, the population composition of L. polyactis showed a trend of miniaturization and younger age (Shan et al., 2017). For another, climate change is regarded as a critical factor that probably enhances or weakens the effects of fishing (Brander, 2007). Liu et al. (2017) found that increasing fishing pressure was the main reason influencing the catch of L. polyactis in the Yellow Sea, and climate change also played a significant role. Above all, the impacts of climate changes on fishery are increasingly important, and more attention should be paid (Liang et al., 2018).
It is helpful to combine the species into groups to study the changing dynamics of these groups in the community (Criales-Hernandez et al., 2006) and understand the internal mechanism of the observed changes (Abdou et al., 2016). In the past eight years, despite the miniaturization of fish, a slight increase in the mean weight proportion of the target fish group was found. L. polyactis and Trichiurus lepturus are important parts of the target fish group. Some previous studies also reported that the catches of these two species increased in recent years (Liu et al., 2017). Not only fishing but also climatic factors like warming seawater temperature may contribute to these results together. However, the mean weight proportion of the non-target fish group unexpectedly underwent a substantial decline in Haizhou Bay. This result was contrary to our hypothesis, supposing that a constant or rising curve may appear. A similar effect was observed in the southeast Atlantic coastal fisheries of the USA (Bacheler and Smart, 2016). Some possible reasons could explain this confusing phenomenon, including bio-invasion (McClenachan et al., 2015), changes in fishers' behavior (Béné and Tewfik, 2001), and global climate change (Bell et al., 2015). With the decline of traditional fish resources in China, crustaceans played an increasingly vital role in the marine fishery (Shen and Heino, 2014). The mean weight proportion of the target crustaceans group showed an apparent upward trend (Fig. 3). Two explanations were put forward. First, with the development of global warming, the annual spawning activities of crustaceans changed (Mao et al., 2019). Increased catching rates of American lobster (Homarus americanus) and blue crab (Callinectes sapidus) were observed because of higher temperature (Jensen et al., 2005; Chang et al., 2010). Second, SMMF is more beneficial to the fast-growing species (Demestre et al., 2008), including many crustaceans like Charybdis bimaculata and P. trituberculatus (Chen, 1991). Scholars usually paid more attention to the target species, while the non-target species' roles in the marine ecosystems cannot be ignored (Bacheler and Smart, 2016). Opposite trends between the target and non-target groups inspired that special studies focusing on the non-target species should be strengthened.
Diversity indices are positively related to the community functions and are used to reveal the changes of the community (Worm et al., 2006). Our results showed that the community compositions of set-net fishery in Haizhou Bay were relatively stable from 2011 to 2018 (Tables 3 & 4). A similar study was carried out in the East China Sea from 2000 to 2007 (Jiang et al., 2009), mean H′ and J′ were 1.97 and 0.51 in April, which was lower than our corresponding results (H′=2.25, J′=0.77) before SMMF. Although two studies were implemented in the two areas, higher indices probably represent a positive status. On the contrary, some bottom trawl surveys in the similar periods reported tremendous declines in fishery resources and diversities in Haizhou Bay because of human disturbance (Su et al., 2015; Zhang et al., 2020). Bottom trawling is regarded as posing severe damages to the marine benthic habitats (Hiddink et al., 2017), while set-net operations are harmless. Habitat destruction may lead to the declines described above. The Shannon diversity after SMMF was lower than before SMMF (Table 3), which could be explained from two aspects. First, the intensity of fishing activities during SMMF decreased to the lowest extent, which promoted the mean weight proportions of the dominant species increase after SMMF. A higher abundance of the dominant species may lower the Shannon diversity (Tsujimoto et al., 2006). Second, seasonality patterns are also important (Hunter et al., 2006; Su et al., 2015).
Increased CPUE after SMMF was observed (Fig. 4), which may benefit from the effects of SMMF on the increasing reproduction rate (Samy-Kamal et al., 2015). However, SMMF does not protect the spawning populations of the all target species since some target species breed before SMMF. The impact of SMMF was limited for O. ocellatus because its spawning time is in the spring (Wang et al., 2015). By contrary, the spawning seasons of T. curvirostris, P. trituberculatus, and O. oratoria were mainly in summer (Chen, 1991). Therefore, SMMF was an effective strategy for the reproduction of these species and the survival of their juvenile. The miniaturization of marine fishery resources has been a great challenge for the sustainable development of China's marine fisheries (Yan et al., 2019). While slightly increasing trends for P. trituberculatus and A. hexanema from 2011 to 2018 were observed (Fig. 3). P. trituberculatus is an important economic crab in China (Dan et al., 2016), which grows fast and has high reproducibility during summer (Hamasaki et al., 2006). Because the recruitment and juvenile stage were highly coincided with SMMF, the CPUE and mean total length of P. trituberculatus increased. As for A. hexanema, two reasons could contribute to the increased total length after SMMF. On the one hand, A. hexanema is a non-target fish with low economic value, and lower fishing intention may protect the stock. On the other hand, 1-year old fish attains sexual maturity, which promoted the growth of the population (Wu and Zhong, 2008).
4.2 Management implicationChina has implemented SMMF for over 27 years, but SMMF is controversial (Jiang et al., 2009; Yan et al., 2019). For the benefit, the fishing intensity was decreased during SMMF, which was advantageous to the reproductive activities of adult fish and the growth of juveniles (Yan et al., 2019). On the opposite, the management cost of SMMF is high, while it cannot fulfill restorative expectations for the fish community (Jiang et al., 2009). This study attempted to evaluate the impact of seasonal closure policy on the fishery and provide suggestions for long-term policy improvement from two aspects.
First, complementary policies matching for SMMF are recommended. Higher abundances of catches could be expected when the annual closure ended. However, with the resumption of fishing activity, CPUE decreased rapidly in several months (Jiang et al., 2009; Yu et al., 2017). What's more, the total length of six species showed downward trends from 2011 to 2018. These results were consistent with the previous studies (Arendse et al., 2007; Yu et al., 2017). During SMMF, strict management was implemented by the Chinese government, which promoted the recovery of fishery resources (Yu et al., 2017). While after SMMF, follow-up measures were deficient. As expected, the fishing stress vengefully increased, and the restored fishery resources were exploited. One of the most vital reasons is fishers' income dropped during SMMF; they wanted to gain more catches after SMMF. Arendse et al. (2007) also pointed out that the closure of the breeding months was not meaningful if annual fishing mortality remains constant for the year. Thus, lacking supporting policies after closure is a critical reason for the poor effects of SMMF.
SMMF is a simple policy itself; however, its complementary measures are systematic and complicated. In order to maintain the effects of SMMF, a comprehensive set of precise and restricted fishery measures must be imposed. Four widely accepted supporting policies are suggested: (1) SMMF needs more enforcement in the fishery management to forbid illegal fishing activity, which is an unrelenting problem for small-scale fisheries worldwide (Nahuelhual et al., 2018); (2) diversified SMMF combined with specific species fishing moratorium and temporary fishing moratorium are suggested (Khan et al., 2018); (3) the fishery institute is suggested to organize the fishing activity of fishers complying with fishery industry rules (Raemaekers et al., 2011); (4) enlarging mesh size, optimizing fishing gear and controlling Total Allowable Catch (TAC) is advisable to be implemented as coordinated measures for SMMF (Karagiannakos, 1996).
Second, with the development of climate change, a new understanding of the biological characteristics like spawning periods for the most species needs to be supplied. The impact of ocean warming on fishery catches has been frequently studied recently (Perry et al., 2005; Cheung et al., 2009; Lam et al., 2020). Sea surface temperature in the Yellow Sea increased by 1.27 ℃ from 1957 to 2006, significantly impacting the Chinese marine fisheries (Liang et al., 2018). One main concern is that warmer sea temperatures may promote the breeding period earlier (Chen et al., 2006). Some studies confirmed this view. Shih et al. (2009) suggested carrying out SMMF from April to June after analyzing the spawning periods of 51 major target fish species in the East China Sea. Feng et al. (2019) advocated that the SMMF should be arranged from April according to the spawning activities of commercial fish in the South China Sea. It is reasonable to determine the closing time based on the breeding seasons of most species. However, the references of fishery biological characteristics in China were almost studied before 2000; some information cannot correspond to the current conditions (Shih et al., 2009). Therefore, comprehensive and systematic fishery surveys should be encouraged.
4.3 LimitationThere were some limitations of our study. First, our results were based on the fishery-dependent data, whose catch and effort information collected by the fishing industry. It is cheaper to obtain a larger number of fishing data, but it is sometimes insufficient for thorough fishery assessment because of unrepresentative data (Dennis et al., 2015). Second, the loss of sampling data prior to the implementation of SMMF made the actual impact of SMMF hard to be evaluated. Which factors caused higher CPUE of the species, SMMF or recruitment, was hard to clarify. The set-net fishery is a kind of small-scale fishery in China. As in most developing countries, data-poor situations pose severe challenges for fishery management (Brooks et al., 2010). Cheng et al. (1999) analyzed the fishing data of economic species in the East China Sea from 1990 to 1998, concluding that SMMF brought positive functions to the fishery. Third, it is confounded whether community changes were caused by SMMF, seasonality (Caddy, 2000), change of fish' behavior (Jupiter et al., 2012), or migration (Hunter et al., 2006). Previous studies showed that the abundance of some species decreased after fishery closure because of the seasonal pattern of biomass (Sardá et al., 1999; Occhipinti-Ambrogi et al., 2005). Not only that, the change of fish' behavior is an important reason for increasing catches after closure (Januchowski-Hartley et al., 2014), which is explained as moving around freely and occupying more habitats due to the absence of fishing activities. Seasonal migrations of migratory species also have important effects on the fishery community, and a study revealed that the fishery landings corresponded closely to the seasonal movement (Hunter et al., 2006). Last, environment information was not available in the present study. However, environmental factors are meaningful to understand the dynamics of the fishery communities (Tang et al., 2018). Despite some drawbacks, we suggested that SMMF is one of the most critical factors affecting the community structure of the set-net fishery. Our results could provide some valuable references for the following fisheries managements and researches.
5 CONCLUSIONThe set-net fishery is a representative small-scale fishery in Haizhou Bay, China, closely related to the livelihood of artisanal fishers. However, the status of fishery resources is declining over the past decades. This study elaborated the temporal variations of set-net fishery communities in Haizhou Bay from 2011 to 2018, and evaluated the effects of SMMF on the communities. Our results elucidated that the mean weight proportion of the target groups kept stable or increased slightly, while the miniaturization of fishery species has emerged these years. The impacts of SMMF were apparent annually in reducing fishing mortality and increasing catches, while many challenges are non-negligible. The responses of species and groups to the SMMF were various, which may be correlated with their biological characteristics and environmental conditions. In order to better maintain the marked effects of SMMF, modifications like assisting with rigorous enforcement and supporting measures are supposed to be implemented. Furthermore, special studies for full understandings of species' spawning periods and investigations aiming at particular species are suggested, which could provide better proposals for policy-makers.
6 DATA AVAILABILITY STATEMENTThe datasets generated and/or analyzed during the current study will be available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENTWe thank all scientific staff and crew members for their assistance in the surveys.
Abdou K, Halouani G, Hattab T, Romdhane M S, Lasram F B R, Le Loc'H F. 2016. Exploring the potential effects of marine protected areas on the ecosystem structure of the Gulf of Gabes using the Ecospace model. Aquatic Living Resources, 29(2): 202.
DOI:10.1051/alr/2016014 |
Anderson M J. 2001. Permutation tests for univariate or multivariate analysis of variance and regression. Canadian Journal of Fisheries and Aquatic Sciences, 58(3): 626-639.
DOI:10.1139/f01-004 |
Arendse C J, Govender A, Branch G M. 2007. Are closed fishing seasons an effective means of increasing reproductive output?: a per-recruit simulation using the limpet Cymbula granatina as a case history. Fisheries Research, 85(1-2): 93-100.
DOI:10.1016/j.fishres.2007.01.001 |
Bacheler N M, Smart T I. 2016. Multi-decadal decline in reef fish abundance and species richness in the southeast USA assessed by standardized trap catches. Marine Biology, 163(2): 26.
DOI:10.1007/s00227-015-2774-x |
Bell R J, Richardson D E, Hare J A, Lynch P D, Fratantoni P S. 2015. Disentangling the effects of climate, abundance, and size on the distribution of marine fish: an example based on four stocks from the northeast US shelf. ICES Journal of Marine Science, 72(5): 1311-1322.
DOI:10.1093/icesjms/fsu217 |
Béné C, Tewfik A. 2001. Fishing effort allocation and fishermen's decision making process in a multi-species small-scale fishery: analysis of the conch and lobster fishery in Turks and Caicos islands. Human Ecology, 29(2): 157-186.
DOI:10.1023/A:1011059830170 |
Brander K M. 2007. The role of growth changes in the decline and recovery of North Atlantic cod stocks since 1970. ICES Journal of Marine Science, 64(2): 211-217.
DOI:10.1093/icesjms/fsl021 |
Brooks E N, Powers J E, Cortes E. 2010. Analytical reference points for age-structured models: application to data-poor fisheries. ICES Journal of Marine Science, 67(1): 165-175.
DOI:10.1093/icesjms/fsp225 |
Caddy J F. 2000. Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas. ICES Journal of Marine Science, 57(3): 628-640.
DOI:10.1006/jmsc.2000.0739 |
Chang J H, Chen Yong, Holland D, Grabowski J. 2010. Estimating spatial distribution of American lobster Homarus americanus using habitat variables. Marine Ecology Progress Series, 420: 145-156.
DOI:10.3354/meps08849 |
Chen D G. 1991. Fishery Ecology in the Yellow Sea and Bohai Sea. China Ocean Press, Beijing, China.
(in Chinese)
|
Chen K S, Crone P, Hsu C C. 2006. Reproductive biology of female pacific bluefin tuna Thunnus orientalis from south-western North Pacific Ocean. Fisheries Science, 72(5): 985-994.
DOI:10.1111/j.1444-2906.2006.01247.x |
Cheng J H, Yan L P, Lin L S, Yu L F, Liu Z F, Zhou Y D, Tang J H, Zhong X M. 1999. Analyses on the fishery ecological effect of summer close season in the East China Sea region. Journal of Fishery Sciences of China, 6(4): 81-85.
(in Chinese with English abstract) |
Cheung W W L, Lam V W Y, Sarmiento J L, Kearney K, Watson R, Pauly D. 2009. Projecting global marine biodiversity impacts under climate change scenarios. Fish and Fisheries, 10(3): 235-251.
DOI:10.1111/j.1467-2979.2008.00315.x |
Clarke K R, Gorley R. 2006. PRIMER v6: user manual/tutorial (Plymouth routines in multivariate ecological research). https://www.primer-e.com.
|
Clarke K R, Warwick R M. 1994. Similarity-based testing for community pattern: the two-way layout with no replication. Marine Biology, 118(1): 167-176.
DOI:10.1007/BF00699231 |
Clarke K R. 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology, 18(1): 117-143.
DOI:10.1111/j.1442-9993.1993.tb00438.x |
Criales-Hernandez M I, Duarte L O, García C B, Manjarrés L. 2006. Ecosystem impacts of the introduction of bycatch reduction devices in a tropical shrimp trawl fishery: insights through simulation. Fisheries Research, 77(3): 333-342.
DOI:10.1016/j.fishres.2005.10.005 |
Dan S, Oshiro M, Ashidate M, Hamasaki K. 2016. Starvation of Artemia in larval rearing water affects post-larval survival and morphology of the swimming crab, Portunus trituberculatus (Brachyura, Portunidae). Aquaculture, 452: 407-415.
DOI:10.1016/j.aquaculture.2015.06.004 |
Davies T D, Baum J K. 2012. Extinction risk and overfishing: reconciling conservation and fisheries perspectives on the status of marine fishes. Scientific Reports, 2: 561.
DOI:10.1038/srep00561 |
Demestre M, de Juan S, Sartor P, Ligas A. 2008. Seasonal closures as a measure of trawling effort control in two Mediterranean trawling grounds: effects on epibenthic communities. Marine Pollution Bulletin, 56(10): 1765-1773.
DOI:10.1016/j.marpolbul.2008.06.004 |
Dennis D, Plagányi É, Van Putten I, Hutton T, Pascoe S. 2015. Cost benefit of fishery-independent surveys: are they worth the money?. Marine Policy, 58: 108-115.
DOI:10.1016/j.marpol.2015.04.016 |
Feng B, Li Z L, Hou G. 2019. On adjustment of closed season in South China Sea. Ocean Development and Management, 36(11): 23-28.
(in Chinese with English abstract) |
Geng Y L, Zhang C L, Han D Y, Ren Y P. 2019. Estimation of growth parameters of Oratosquilla oratoria based on fishery-independent and -dependent data collected from the coast of Shandong Province. Journal of Fishery Sciences of China, 26(4): 756-764.
(in Chinese with English abstract) |
Hamasaki K, Fukunaga K, Kitada S. 2006. Batch fecundity of the swimming crab Portunus trituberculatus (Brachyura: Portunidae). Aquaculture, 253(1-4): 359-365.
DOI:10.1016/j.aquaculture.2005.08.002 |
Hiddink J G, Jennings S, Sciberras M, Szostek C L, Hughes K M, Ellis N, Rijnsdorp A D, McConnaughey R A, Mazor T, Hilborn R, Collie J S, Pitcher C R, Amoroso R O, Parma A M, Suuronen P, Kaiser M J. 2017. Global analysis of depletion and recovery of seabed biota after bottom trawling disturbance. Proceedings of the National Academy of Sciences of the United States of America, 114(31): 8301-8306.
DOI:10.1073/pnas.1618858114 |
Hunter E, Berry F, Buckley A A, Stewart C, Metcalfe J D. 2006. Seasonal migration of thornback rays and implications for closure management. Journal of Applied Ecology, 43(4): 710-720.
DOI:10.1111/j.1365-2664.2006.01194.x |
Jackson J B C, Kirby M X, Berger W H, Bjorndal K A, Botsford L W, Bourque B J, Bradbury R H, Cooke R, Erlandson J, Estes J A, Hughes T P, Kidwell S, Lange C B, Lenihan H S, Pandolfi J M, Peterson C H, Steneck R S, Tegner M J, Warner R R. 2001. Historical overfishing and the recent collapse of coastal ecosystems. Science, 293(5530): 629-637.
DOI:10.1126/science.1059199 |
Januchowski-Hartley F A, Cinner J E, Graham N A J. 2014. Fishery benefits from behavioural modification of fishes in periodically harvested fisheries closures. Aquatic Conservation: Marine and Freshwater Ecosystems, 24(6): 777-790.
DOI:10.1002/aqc.2388 |
Jensen O P, Seppelt R, Miller T J, Bauer L J. 2005. Winter distribution of blue crab Callinectes sapidus in Chesapeake Bay: application and cross-validation of a two-stage generalized additive model. Marine Ecology Progress Series, 299: 239-255.
DOI:10.3354/meps299239 |
Jiang Y Z, Cheng J H, Li S F. 2009. Temporal changes in the fish community resulting from a summer fishing moratorium in the northern East China Sea. Marine Ecology Progress Series, 387: 265-273.
DOI:10.3354/meps08078 |
Jin Y, Chen X J, Chen P. 2018. Preliminary study of biological characteristics of Octopus ocellatus in the East China Sea and the Yellow Sea in Autumn. Journal of Guangdong Ocean University, 38(5): 15-20.
(in Chinese with English abstract) |
Jupiter S D, Weeks R, Jenkins A P, Egli D P, Cakacaka A. 2012. Effects of a single intensive harvest event on fish populations inside a customary marine closure. Coral Reefs, 31(2): 321-334.
DOI:10.1007/s00338-012-0888-x |
Karagiannakos A. 1996. Total allowable catch (TAC) and quota management system in the European Union. Marine Policy, 20(3): 235-248.
DOI:10.1016/0308-597X(96)86986-7 |
Khan A M A, Gray T S, Mill A C, Polunin N V C. 2018. Impact of a fishing moratorium on a tuna pole-and-line fishery in eastern Indonesia. Marine Policy, 94: 143-149.
DOI:10.1016/j.marpol.2018.05.014 |
Kuparinen A, Merilä J. 2007. Detecting and managing fisheries-induced evolution. Trends in Ecology & Evolution, 22(12): 652-659.
DOI:10.1016/j.tree.2007.08.011 |
Lam V W Y, Allison E H, Bell J D, Blythe J, Cheung W W L, Frölicher T L, Gasalla M A, Sumaila U R. 2020. Climate change, tropical fisheries and prospects for sustainable development. Nature Reviews Earth & Environment, 1(9): 440-454.
DOI:10.1038/s43017-020-0071-9 |
Liang C, Xian W W, Pauly D. 2018. Impacts of ocean warming on China's fisheries catches: an application of "Mean Temperature of the Catch" concept. Frontiers in Marine Science, 5: 26.
DOI:10.3389/fmars.2018.00026 |
Liu X X, Wang J, Xu B D, Xue Y, Ren Y P. 2017. Impacts of fishing pressure and climate change on catches of small yellow croaker in the Yellow Sea and the Bohai Sea. Periodical of Ocean University of China, 47(8): 58-64.
(in Chinese with English abstract) |
Ludwig J A, Reynolds J F. 1988. Statistical Ecology: a Primer on Methods and Computing. Wiley, New York.
|
Mao X Y, Guo X Y, Wang Y C, Takayama K. 2019. Influences of global warming on the larval survival and transport of snow crab (Chionoecetes opilio) in the Sea of Japan. Sustainability, 11(8): 2198.
DOI:10.3390/su11082198 |
McClanahan T R. 2014. Recovery of functional groups and trophic relationships in tropical fisheries closures. Marine Ecology Progress Series, 497: 13-23.
DOI:10.3354/meps10605 |
McClenachan L, O'Connor G, Reynolds T. 2015. Adaptive capacity of co-management systems in the face of environmental change: the soft-shell clam fishery and invasive green crabs in Maine. Marine Policy, 52: 26-32.
DOI:10.1016/j.marpol.2014.10.023 |
Nahuelhual L, Saavedra G, Blanco G, Wesselink E, Campos G, Vergara X. 2018. On super fishers and black capture: images of illegal fishing in artisanal fisheries of southern Chile. Marine Policy, 95: 36-45.
DOI:10.1016/j.marpol.2018.06.020 |
Occhipinti-Ambrogi A, Savini D, Forni G. 2005. Macrobenthos community structural changes off Cesenatico Coast (Emilia Romagna, Northern Adriatic), a six-year monitoring programme. Science of the Total Environment, 353(1-3): 317-328.
DOI:10.1016/j.scitotenv.2005.09.021 |
Pang Z W, Xu B D, Zan X X, Ren Y P. 2015. Shrimp community structure and its relationships with environmental factors in Haizhou Bay and adjacent waters in spring. Acta Ecologica Sinica, 35(6): 191-195.
DOI:10.1016/j.chnaes.2015.09.005 |
Pei R D, Gao C X, Tian S Q, Ma Q Y, Fan Q S. 2021. Heterogeneity of growth and maturity of Larimichthys polyactis in the offshore waters of southern Zhejiang, China. Chinese Journal of Applied Ecology, 32(1): 333-341.
(in Chinese with English abstract) |
Perry A L, Low P J, Ellis J R, Reynolds J D. 2005. Climate change and distribution shifts in marine fishes. Science, 308(5730): 1912-1915.
DOI:10.1126/science.1111322 |
Pielou E C. 1966. Species-diversity and pattern-diversity in the study of ecological succession. Journal of Theoretical Biology, 10(2): 370-383.
DOI:10.1016/0022-5193(66)90133-0 |
Raemaekers S, Hauck M, Bürgener M, Mackenzie A, Maharaj G, Plagányi É E, Britz P J. 2011. Review of the causes of the rise of the illegal south African abalone fishery and consequent closure of the rights-based fishery. Ocean & Coastal Management, 54(6): 433-445.
DOI:10.1016/j.ocecoaman.2011.02.001 |
Rice J C. 2000. Evaluating fishery impacts using metrics of community structure. ICES Journal of Marine Science, 57(3): 682-688.
DOI:10.1006/jmsc.2000.0735 |
Samy-Kamal M, Forcada A, Lizaso J L S. 2015. Effects of seasonal closures in a multi-specific fishery. Fisheries Research, 172: 303-317.
DOI:10.1016/j.fishres.2015.07.027 |
Sardá R, Pinedo S, Martin D. 1999. Seasonal dynamics of macroinfaunal key species inhabiting shallow soft-bottoms in the bay of Blanes (NW Mediterranean). Acta Oecologica, 20(4): 315-326.
DOI:10.1016/S1146-609X(99)00135-6 |
Shan X J, Li X S, Yang T, Sharifuzzaman S M, Zhang G Z, Jin X S, Dai F Q. 2017. Biological responses of small yellow croaker (Larimichthys polyactis) to multiple stressors: a case study in the Yellow Sea, China. Acta Oceanologica Sinica, 36(10): 39-47.
DOI:10.1007/s13131-017-1091-2 |
Shen G M, Heino M. 2014. An overview of marine fisheries management in China. Marine Policy, 44: 265-272.
DOI:10.1016/j.marpol.2013.09.012 |
Shih N T, Cai Y H, Ni I H. 2009. A concept to protect fisheries recruits by seasonal closure during spawning periods for commercial fishes off Taiwan and the East China Sea. Journal of Applied Ichthyology, 25(6): 676-685.
DOI:10.1111/j.1439-0426.2009.01328.x |
Simpson E H. 1949. Measurement of diversity. Nature, 163(4148): 688.
DOI:10.1038/163688a0 |
Sowman M. 2006. Subsistence and small-scale fisheries in South Africa: a ten-year review. Marine Policy, 30(1): 60-73.
DOI:10.1016/j.marpol.2005.06.014 |
Su W, Xue Y, Zhang C L, Ren Y P. 2015. Spatio-seasonal patterns of fish diversity, Haizhou Bay, China. Chinese Journal of Oceanology and Limnology, 33(1): 121-134.
DOI:10.1007/s00343-015-3311-y |
Sun Z Z, Zhou J, Zhao Z L, Xu Y F, Gao W B, Zhuang S, Chen B, Wang J, Yan W, Wang C H, Meng W D. 2012. The fishing structure of the Yellow Sea and Bohai Sea. Marine Science, 36(6): 44-53.
(in Chinese with English abstract) |
Suuronen P, Chopin F, Glass C, Løkkeborg S, Matsushita Y, Queirolo D, Rihan D. 2012. Low impact and fuel efficient fishing-looking beyond the horizon. Fisheries Research, 119-120: 135-146.
DOI:10.1016/j.fishres.2011.12.009 |
Tang Y L, Ma S Y, Liu C D, Wang X M, Cheng S S. 2018. Influence of spatial-temporal and environmental factors on Larimichthys polyactis, Octopus variabilis, and species aggregated Set-net CPUEs in Haizhou Bay, China. Journal of Ocean University of China, 17(4): 973-982.
DOI:10.1007/s11802-018-3659-7 |
Tang Y L, Qi G R, Wang X, Tian F, Wan R. 2014. Catch composition of the Set-Net along the coastal water and resource utilization analysis of Haizhou Bay. Periodical of Ocean University of China, 44(7): 29-38.
(in Chinese with English abstract) |
Tsujimoto A, Nomura R, Yasuhara M, Yamazaki H, Yoshikawa S. 2006. Impact of eutrophication on shallow marine benthic foraminifers over the last 150 years in Osaka Bay, Japan. Marine Micropaleontology, 60(4): 258-268.
DOI:10.1016/j.marmicro.2006.06.001 |
Wang W J, Dong G, Yang J M, Zheng X D, Wei X M, Sun G H. 2015. The development process and seasonal changes of the gonad in Octopus ocellatus gray off the coast of Qingdao, northeast China. Fisheries Science, 81(2): 309-319.
DOI:10.1007/s12562-014-0846-3 |
Worm B, Barbier E B, Beaumont N, Duffy J E, Folke C, Halpern B S, Jackson J B C, Lotze H K, Micheli F, Palumbi S R, Sala E, Selkoe K A, Stachowicz J J, Watson R. 2006. Impacts of biodiversity loss on ocean ecosystem services. Science, 314(5800): 787-790.
DOI:10.1126/science.1132294 |
Wu H L, Zhong J S. 2008. Fauna Sinica, Teleosta, Perciformes, Gobioodei. Science Press, Beijing, China.
(in Chinese)
|
Xu B Q, Chen W, Wang T T, Yang Y Y, Li F, Liu S D. 2021. Resource status and distribution characteristics of Portunus trituberculatus before the end of summer fishing moratorium in Laizhou Bay. Journal of Fisheries of China, 45(4): 543-551.
(in Chinese with English abstract) |
Yan L P, Ling J Z, Li J S, Lin L S, Cheng J H. 2006. Simulative analysis on results of summer closed fishing in the East China Sea with Ricker population dynamic pool model. Journal of Fishery Sciences of China, 13(1): 85-91.
(in Chinese with English abstract) |
Yan L P, Liu Z L, Jin Y, Cheng J H. 2019. Effects of prolonging summer fishing moratorium in the East China Sea on the increment of fishery resources. Marine Fisheries, 41(5): 513-519.
(in Chinese with English abstract) |
Yan L, Tan Y G, Yang B Z, Zhang P, Li J, Yang L. 2016. Comparison on resources community of stow-net fishery before and after fishing off season in Huangmaohai Estuary. South China Fisheries Science, 12(6): 1-8.
(in Chinese with English abstract) |
Yan Y Y, Lu C C. 2020. Suggestions on adjusting the summer fishing moratorium. Journal of Zhejiang Ocean University (Natural Science), 39(2): 163-166, 179.
(in Chinese with English abstract) |
Yang C X, Zhu Y G, Wan R, Zhang J B. 2011. Application of fishermen's organizations in small-scale fisheries management. Chinese Fisheries Economics, 29(2): 63-68.
(in Chinese with English abstract) |
Yu J, Hu Q W, Yuan H R, Chen P M. 2018. Effect assessment of summer fishing moratorium in Daya Bay based on remote sensing data. South China Fisheries Science, 14(3): 1-9.
(in Chinese with English abstract) |
Yu J, Hu Q W, Yuan H R, Tong F, Chen P M, Mao J W. 2017. Effects assessment of summer fishing moratorium in Daya Bay in the Northern South China Sea. Journal of Geoscience and Environment Protection, 5(7): 96-108.
DOI:10.4236/gep.2017.57010 |
Yue D D, Wang L M, Xiong M S, Xiao L, Zhang H Y. 2016. Improving the marine summer closed fishing season system: A case study of the practice of Zhejiang Province and the East China Sea. Research of Agricultural Modernization, 37(2): 337-344.
(in Chinese with English abstract) |
Zhang Y L, Yu H M, Yu H Q, Xu B D, Zhang C L, Ren Y P, Xue Y, Xu L L. 2020. Optimization of environmental variables in habitat suitability modeling for mantis shrimp Oratosquilla oratoria in the Haizhou bay and adjacent waters. Acta Oceanologica Sinica, 39(6): 36-47.
DOI:10.1007/s13131-020-1546-8 |
Zou J W, Wang Q Z, Huang J X, Yang B H, Pan P. 2016. Evaluation of fishing moratorium on fishing grounds in the northern continental shelf of the South China Sea in summer. Fisheries Science & Technology Information, 43(6): 318-323.
(in Chinese) |