 2022, Vol. 40
2022, Vol. 40Institute of Oceanology, Chinese Academy of Sciences
Article Information
- WANG Junjian, LI Chaolun, YANG Guang, TAO Zhencheng, WANG Yanqing, XIAN Haochen
- Dietary preferences and potential ecological impact on the zooplankton community of Nemopilema nomurai based on stable isotope and fatty acid analyses
- Journal of Oceanology and Limnology, 40(3): 1085-1096
- http://dx.doi.org/10.1007/s00343-021-1024-y
Article History
- Received Jan. 20, 2021
- accepted in principle May. 6, 2021
- accepted for publication Jun. 24, 2021




2 Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China;
3 North China Sea Marine Forecasting Center of State Oceanic Administration, Qingdao 266100, China;
4 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China;
5 University of Chinese Academy of Sciences, Beijing 100049, China;
6 Department of Engineering and Technology Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
Blooms of the giant jellyfish Nemopilema nomurai have occurred frequently in the Yellow Sea since 2000 (Zhang et al., 2012; Uye, 2014; Yoon et al., 2014; Sun et al., 2015). During late summer, the biomass of N. nomurai in the Yellow Sea can reach millions of tons (Sun et al., 2015). Explosive blooms of N. nomurai can disrupt the fish-dominated trophic structure, transforming it into one dominated by jellyfish (Kawahara et al., 2006; Uye, 2008, 2011, 2014). Fisheries may also be threatened (Ding and Cheng, 2005; Wu et al., 2008; Zhang et al., 2012)
The metephyrae of N. nomurai can be found every spring in the Yellow Sea. Before blooming in late summer, N. nomurai needs abundant food to support rapid growth. Thus, sufficient food would provide the energy base for blooming. Various micro- and meso-zooplankton, such as bacteria, ciliates, copepods, fish eggs, and juvenile fish, may be food sources for N. nomurai (Uye, 2008). However, specific nutrition requirements of N. nomurai are not well understood. Additionally, large numbers of N. nomurai may exert a severe grazing impact on its prey (Uye, 2014). Understanding the dietary composition of N. nomurai under natural conditions is essential to revealing the nutritional drivers and ecological consequences of its blooms.
Nemopilema nomurai is a species of Rhizostomeae jellyfish. Unlike the other common Scyphozoa jellyfish, the central mouth of N. nomurai closes when an ephyra becomes medusae. The terminal pore located on its oral arm then becomes its main feeding organ. While N. nomurai is usually unable to consume zooplankton larger than its terminal pore (Lee et al., 2008), Pitt et al. (2009) indicated that large zooplankton might also be a major food source for Rhizostomeae medusae. The typical prey size of N. nomurai remains to be verified. Moreover, the shapes of its potential food sources (e.g., cylindric copepods and spherical fish eggs) differ, which may affect the consumption. Therefore, the dietary composition of N. nomurai under natural conditions shall be further investigated.
Compared with the other jellyfish, N. nomurai is relatively large. The bell diameter may reach 2 m and the wet weight up to 200 kg (Omori and Kitamura, 2004). The large size and fragile body of N. nomurai make capture and cultivation difficult, thus culture experiments for studying feeding mechanisms are not practical for this species. Gut content analyses is well used in research of diet of jellyfish. For example, gut content analyses revealed that Aurelia aurita consumed mainly copepods predator (Ishii and Tanaka, 2001; Lo and Chen, 2008); Aurelia solida fed on Calanoid copepods, mollusc larvae, and larvaceans (Gueroun et al., 2020); Pelagia noctiluca ate fish eggs and larvae (Tilves et al., 2016). Uye (2008) summarized that among the food spectra of N. nomurai, most consisted of micro- and meso-zooplankton. However, some issues remain. For examples, food intensive effects during sample collection may affect the results, and food quantity cannot be measured.
Biomarker methods based on stable isotopes and fatty acids (FAs) may be beneficial for studies on the diet of N. nomurai. In predators, heavier stable isotopes may be accumulated more than lighter ones, making stable isotopes such as 13C/12C and 15N/14N effective tools for understanding the food sources and trophic positions of organisms in the food web (Post, 2002). FA biomarkers may also be useful for studying the dietary characteristics of consumers because certain FAs in consumers can stay unchanged or change in specific ways (Pitt et al., 2009). Compared with gut content analyses, they are advantageous for quantifying food composition and imparting information on the long-term diet of the consumer (Kling et al., 1992; Cabana and Rasmussen, 1996; Teuber et al., 2014; Henschke et al., 2015; Yang et al., 2016). These methods have been used to investigate the diets of creatures in the marine food web (El-Sabaawi et al., 2009; Allan et al., 2010; Connelly et al., 2014; Wang et al., 2015). In recent years, stable isotope and FA biomarkers have been used successfully in studies on the dietary features of jellyfish. Using these markers, it was found that the trophic niche of Pelagia noctiluca overlapped highly with fish larvae (Tilves et al., 2018), and microzooplankton were important prey of this jellyfish (Milisenda et al., 2018). FA biomarkers showed that the availability of living plankton or suspended detritus could affect the diet of P. noctiluca (Milisenda et al., 2018). Cui et al. (2012) found that zooplankton were important in the diets of large jellyfish in the Yellow Sea. These methods are feasible for diet study of N. nomurai.
To clarify the information on the long-term diet of N. nomurai is helpful to study the relationship between N. nomurai and zooplanktons. Because of the special feeding organ of N. nomurai, we hypothesized that small creatures or organic matter contributed more to the food of N. nomurai than the large ones. To prove this hypothesis, we investigated the FAs and two stable isotopes (13C and 15N) of N. nomurai collected in August 2016, then analyzed the dietary composition of N. nomurai. The impact of N. nomurai on the zooplankton community was also evaluated.
2 MATERIAL AND METHOD 2.1 Sample collectionDuring a survey conducted in the Yellow Sea (122°E–127°E, 30°N–38°N) from the research vessel (R/V) Beidou in August 2016, samples of N. nomurai, zooplankton, and particulate organic matter (POM) were taken at 11 stations (Fig. 1). To collect N. nomurai samples, bottom trawl nets (length 83.2 m, open circumference 167.2 m, height 7 m, width 22 m, and mesh size 20 cm) were deployed at each station.Because gelatinous zooplankton are usually broken during towing of the trawl net, we kept the towing speed below 3 kt to avoid sample loss. The towing time was 1 h. Subsequently, N. nomurai measuring either > 80 cm or 20–30 cm in diameter were selected to test for differences in dietary composition between large and small individuals. Large N. nomurai (LNN) was used to refer to specimens > 80 cm in diameter, whereas small N. nomurai (SNN) to those 20–30-cm diameter. Parts of the umbrella of N. nomurai were cut and rinsed carefully with distilled water. Samples were then stored in liquid nitrogen for isotope and FA analyses. Zooplankton larger than 505 μm were collected using a maxi-zooplankton net (mesh size 505 μm; diameter 50 cm), while zooplankton with size 160–505 μm were collected with a midi-zooplankton net (mesh size 160 μm; diameter 31.6 cm) (Sun et al., 2010). At each station, duplicate hauls were taken. The first was preserved in a 5% solution of buffered formalin for later counting in the laboratory. The second was picked alive and sorted to the species level under a stereomicroscope. The samples were then rinsed with filtered seawater and distilled water in turn, concentrated on GF/F Whatman filters (diameter=25 mm), and immediately frozen in liquid nitrogen for further FA and stable isotope analyses. Water samples (50 L) were collected to gather POM. After prescreening through a 200-μm sieve to remove large species, POM samples were also filtered onto GF/F filters.

|
| Fig.1 Location of the sampling stations These contour lines are isobaths which were measured in meter. |
The calculation of N. nomurai abundance followed that of Zhang et al. (2012). The following equations were used:
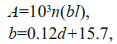
where A is N. nomurai abundance (inds./km2); n is the number of N. nomurai in one catch; and l is the integral distance of a trawl (km), read from a Simrad EK500 echo sounder integral system. The second formula was determined by Tang et al. (2006), where d is the average water depth (m) at each station.
Zooplankton samples were counted under a stereomicroscope. The dominance index (Y) was used to identify dominant species (Odum, 1959). The formula was as follows:
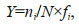
where ni is the species abundance, N is the total abundance of all zooplankton species, and fi is the occurring frequency of species at all stations.Y > 0.02 indicated that the species was dominant (Odum, 1959).
The biovolume of zooplankton was quantified using the semi-automatic ZooScan system (Hydroptic, France). Zooplankton samples were scanned and digitized at a resolution of 2 400 dpi. The major and minor axes of each object were then automatically identified (Grosjean et al., 2004; Gorsky et al., 2010). Zooplankton volume was equal to the ellipsoidal volume. Zooplankton biovolume (mm3/L) was calculated using the following formula:
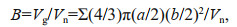
where B is the zooplankton biovolume (mm3/L), Vg is total volume of every organism in each size class (i.e., 200–1 000 μm, 1 000–5 000 μm, and > 5 000 μm), a and b are the major and minor axes (mm) of the zooplankton individuals, respectively, and Vn is the net volume.
FAs were extracted from the samples following the methods previously described by Folch et al. (1957), Kattner and Frickle (1986), and El-Sabaawi et al. (2009). After freeze-drying all the frozen sample, 1.5–2.0 mg of each sample was preserved for the stable isotope analysis. Twenty mg of jellyfish samples and 5 mg of zooplankton and POM samples were weighed out for FAs analyses. Then, the C19:0 saturated FAs were added to the samples as an internal standard. Afterwards, the sample was sonicated and centrifuged (2 400 r/min) in a mixture of chloroform, methanol, and water (ratio 8:4:3) three times to pool the lipid to the organic layers. The layers were extracted and mixed with 0.2-mL methanol containing 3% sulfuric acid at 80 ℃ in a water bath for 4 h. The FAs methyl esters (FAMEs) were then prepared. After cooling, hexane was added to extract the FAMEs. An Agilent 7890A gas chromatograph (Agilent Technologies, Inc., Santa Clara, CA, USA) was used to quantify the FAs. FAs were identified by gas chromatography-mass spectrometry (GC/MS). According to the internal standard method, FAs were calculated quantitatively. The FAs were then expressed as the % of total FAs.
To assess the degree of similarity between FA samples, a cluster analysis was conducted using the PRIMER statistical package after converting the data into similarity triangular matrices with a Bray-Curtis resemblance measure (Bray and Curtis, 1957).
Stable isotopes were determined using a Thermo Delta Ⅳ isotope ratio mass spectrometer (Thermo Fisher Scientific, Inc., Bremen, Germany). Stable isotopic values were expressed in the conventional δ notation and calculated using the following equations:
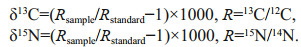
The standard reference materials were Pee Dee Belemnite (PDB) and atmospheric N2, respectively.
Lipids were not removed prior to stable isotope analysis (Schukat et al., 2014). Because δ13C values may be affected by lipid content leading to a depleted δ13C result (DeNiro and Epstein, 1978; Post, 2007), the lipid correction model for aquatic invertebrates described in Logan et al. (2008) was used. The model was as follows:
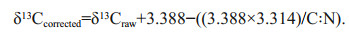
The mixed model Stable Isotope Analysis in R (SIAR) of Parnell et al. (2010) was used to calculate the contribution of different sources to the diet of N.nomurai. To reduce the number of sources substituted into the model and avoid deviation caused by some low-abundance foods, four particle-size groups—i.e., < 200 μm (small organisms), 200–1 000μm (small copepods), 1 000–5 000 μm (large copepods), and > 5 000 μm (giant crustaceans)—were defined according to Sun et al. (2010) and Li et al. (2013) prior to analyses. POM, Oithona similis, Calanus sinicus, and Euphausia pacifica were selected as representative sources of the small organism, small copepod, large copepod, and giant crustacean size groups, respectively. The latter three species were chosen because they were the most abundant species in each group. The trophic enrichment factors used were 0.4‰±1.3‰ (for δ13C) and 3.4‰±1.0‰ (for δ15N) (Post, 2002).
Statistical analyses were conducted using Microsoft Excel 2016 and SPSS 15.0. Primer 6.0 was used to execute the cluster analysis. The isotope mixing models in SIAR package were used to evaluate the contribution of sources.
3 RESULT 3.1 Distribution of N. nomurai and zooplankton in the Yellow SeaNemopilema nomurai was found at all sampling stations, with abundances ranging from 59.23 to 1 651.21 inds./km2 (Fig. 2). The highest and lowest abundances were found at stations A5 and B8, respectively. The average abundance was 401.01 inds./km2. The biovolume of zooplankton varied from 0.38 to 68.59 mm3/L (Fig. 2). A negative correlation was found between N. nomurai and the biovolume of zooplankton (RELATE, R=-0.24, P < 0.05).

|
| Fig.2 Abundance of N. nomurai (inds./km2) (a) and biovolume composition of zooplankton in each size (b) in the Yellow Sea during August, 2016 These contour lines are isobaths which were measured in meter. |
Twenty-six species of zooplankton adults and ten species of zooplankton larvae were identified. O. similis and C. sinicus were the dominant species in the study area (Y=0.22 and Y=0.02, respectively).O. similis was the largest contributor to the abundance of small copepods at most stations (Table 1). The average abundance of O. similis was 550.70 inds./m3, and its maximum abundance occurred at station A2 (1 678.05 inds./m3). C. sinicus was the main contributor to the abundance of large copepods, with an average abundance of 67.13 inds./m3. Its highest abundance of 131.79 inds./m3 was found at station B2 (Table 1). The most abundant giant crustacean was E. pacifica.

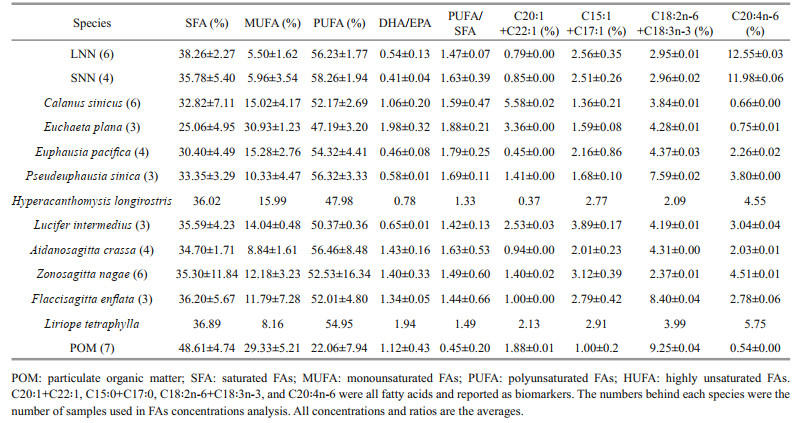
The FAs in the N. nomurai samples were mainly composed of saturated fatty acids (SFAs) and polyunsaturated fatty acids (PUFAs). These two groups contributed > 94% of the total FAs (Table 2). There were no significant differences between SNN and LNN in terms of SFA or PUFA content (ANOVA, P > 0.05). Eicosapentaenoic acid (EPA) (C20:5n-3), arachidonic acid (C20:4n-6), and docosahexaenoic acid (DHA) (C22:6n-3) were the dominant PUFAs in N. nomurai. The average proportion of C20:4n-6 was approximately 12% in both LNN and SNN. C20:4n-6 could be used to detect detritus-based food sources (Dalsgaard et al., 2003). DHA (C22:6n-3) was slightly lower than C20:4n-6 at 11.4% and 10.4% for LNN and SNN, respectively. The FA biomarker DHA/EPA that could distinguish dinoflagellate- and diatom-based diets (Budge and Parrish, 1998) was 0.54% and 0.41% in LNN and SNN, respectively (Table 2). The relative concentrations of C18:2n-6+C18:3n-3, which represented the contribution of terrestrial nutrition (Dalsgaard et al., 2003) were 2.95% in LNN and 2.96% in SNN. The relative concentrations of the bacterial food biomarker C15:0+C17:0 (Kaneda, 1991) was 2.56% and 2.51% in LNN and SNN, respectively. No significant differences between LNN and SNN were found for any of the biomarkers mentioned above (ANOVA, all P values > 0.05).
|
Nemopilema nomurai and zooplankton species were separated into three groups according to cluster analysis (Fig. 3): Groups A and B represented the majority of large copepods, including C. sinicus and Euchaeta plana. Group C included LNN, SNN, and several other species. LNN and SNN sampled at the same station shared more than 90% similarity. Group C was subdivided into two smaller groups with 65% similarity: Group CI included all mysids, arrow worms, and some copepods, and Group CII included N. nomurai, E. pacifica, and Lucifer intermedius. The Group CII could also be divided into two parts with 72% similarity.

|
| Fig.3 Average-neighbor cluster analysis based on a Bray-Curtis dissimilarity matrix of the raw FAs data of species collected in August, 2016 Data are expressed as the percentage of total FAs. |
The LNN and SNN were not significantly different in terms of δ13C and δ15N (ANOVA, P > 0.05) (Fig. 4). They shared a similar δ13C value (-19.84‰±1.4‰ for LNN and -19.91‰±1.3‰ for SNN). The trophic levels of both size classes of N. nomurai were also similar, as indicated by similar δ15N values (5.82‰±1.4‰ for LNN and 5.61‰±1.0‰ for SNN).N. nomurai, O. similis, and E. pacifica were characterized by similar values of δ15N (6.74‰±1.0‰ and 6.49‰±1.0‰ for O. similis and E. pacifica, respectively).

|
| Fig.4 Stable isotope bi-plot based on species collected from the Yellow Sea LNN: large N.nomurai; SNN: small N. nomurai; POM: particulate organic matter. Each point on the graph represents the mean value of 2-8 individuals of a particular species. Data are in mean±SD. |
The average δ13C of POM was -20.62‰±2.0‰. N. nomurai, O. similis, E. pacifica, C. sinicus, Themisto gracilipes, and erichthus larvae had similar δ13C values to that of POM (from -21.89‰±1.3‰ to-19.47‰±0.8‰). The other zooplankton were enriched in δ13C with values ranging from-18.66‰±1.3‰ to 17.35‰±1.0‰.
The δ15N value of POM was 3.51‰±0.93‰, which was lower than that of all the zooplankton. Fish larvae had the highest δ15N value (12.72‰±0.3‰) among all the zooplankton. Zonosagitta nagae was slightly lower than fish larvae with δ15N at 11.56‰±0.6‰. Thermisto gracilipes, Aidanosagitta crassa, erichthus larvae, L. intermedius, and Pseudeuphausia sinica occupied a similar trophic level as indicated by similar values of δ15N (range: 7.56‰ to 8.32‰) (Fig. 4).
SIAR analysis revealed that the contribution of prey larger than 1 000 μm to the diet of N. nomurai was relatively low. The relatively large zooplankton species, C. sinicus and E. pacifica, were consumed less than smaller species by N. nomurai, comprising 16% and 13% of its diet, respectively. Instead, N. nomurai tended to consume smaller food particles and prey such as POM and O. similis, which comprised 38% and 33% of the N. nomurai diet, respectively (Fig. 5). The dietary composition of SNN was similar to that of LNN.

|
| Fig.5 Results of isotope mixing models in SIAR (95%, 75%, and 50% credibility intervals (shown by different gray scale)) of the contribution ratio of sources to Nemopilema nomurai diet POM: particulate organic matter. |
A previous study indicates that N. nomurai began to appear in the Yellow Sea in late May and that its population increased rapidly from June to August, peaking in September or October (Sun et al., 2015). The sampling period for this study was August 2016, a period of rapid growth for N. nomurai in the Yellow Sea. Stable isotope and FA analyses may reflect nutrient absorption during this period.
The average abundance of N. nomurai in the Yellow Sea in August 2016 was 401 inds./km2, which was close to the 689 inds./km2 observed in August 2013 (Sun et al., 2015). Because 2013 was a mid-bloom year for N. nomurai (Sun et al., 2015), we assumed that the sampling period used in this study was also during a mid-bloom year for N. nomurai. From 2006 to 2013, the average abundances of N. nomurai collected by bottom trawl from the Yellow Sea ranged from 34 inds./km2 to 4 951 inds./km2 (Sun et al., 2015). In this study, the abundance of N. nomurai was also within this range. Therefore, the N. nomurai collected here were in a regular state, and our results may be representative of the typical diet for most N. nomurai in the Yellow Sea.
The results of the stable isotope analysis show that POM and small copepods (dominated by O. similis) measuring less than 1 mm comprised over 60% of N. nomurai diet (Fig. 5). Thus, despite its large body size, N. nomurai still preys predominantly on small plankton and organic particles, and this is mainly determined by the structure of N. nomurai feeding organ and its predation method. N. nomurai is a Rhizostomeae jellyfish, and its preys are engulfed into the terminal pore and travel through the canal system to the gut. The diameter of the terminal pore directly determines the size of the food particle that can be ingested by N. nomurai, and regardless of body size, its terminal pore diameter is always about 1 mm (Lee et al., 2008). Thus, it is theoretically difficult for N. nomurai to engulf particles exceeding 1 mm in diameter. Some larger but slender zooplankton may also be preyed upon by Rhizostomeae (Pitt et al., 2009). Therefore, we considered large copepods and krill sizes greater than 1 mm to be potential prey items for N. nomurai. The SIAR results show that although a high biomass of zooplankton larger than 1 mm was observed during the survey period (Fig. 2), and their contribution to N. nomurai diet was much lower than that of small copepods and POM (Fig. 5). According to the studies of Chinese researchers, the POM in the South Yellow Sea was mainly composed by resuspension and river input detritus, as well as phytoplankton and microzooplanktons (Qin et al., 1988, 1989; Liu et al., 2010; Hu et al., 2017).Therefore, N. nomurai blooms may impose relatively high feeding pressure on small creatures which include small copepods, microzooplankton, and phytoplankton.
The FA analysis results showed that the content of FAs C18:2n-6+C18:3n-3 in POM was higher than in all the other zooplankton (Table 2). In previous studies, C18:2n-6+C18:3n-3 was used as a biomarker for terrestrial material. The threshold to distinguish between terrestrial and marine sources was considered 2.5% (Parrish et al., 1995; Budge and Parrish, 1998). When the C18:2n-6+C18:3n-3 content in the FAs of a species is higher than 2.5%, its diet likely contains substances of terrestrial origin (Budge and Parrish, 1998). Therefore, given that the C18:2n-6+C18:3n-3 content in POM in this survey was much higher than 2.5%, it may be that the POM in the Yellow Sea contains large quantities of terrestrial components. Considering POM's large contribution to the diet of N. nomurai, terrestrial sources of nutrition should have taken a certain contribution to N. nomurai diet. Therefore, terrestrial inputs may promote blooms of N. nomurai.
4.2 Impact of N. nomurai on zooplankton communityThe FA clustering results showed that the FA composition of N. nomurai was similar to that of the krill E. pacifica (Fig. 3). Past studies indicate that as a food source of N. nomurai, most small plankton can also be potential food sources for E. pacifica (Ohman, 1984; Dilling et al., 1998; Nakagawa et al., 2004; Bargu et al., 2006). Because the size of the terminal pore makes it difficult for N. nomurai to feed on organisms larger than 5 000 μm, including krill, the only explanation for the similarities between N. nomurai and krill in terms of FA composition is that they had ingested similar food sources. Having a similar diet could lead to competition for food resources. Therefore, N. nomurai blooms could have a negative impact on E. pacifica population growth.E. pacifica is the main food for many marine vertebrates, including fish and seabirds (Brinton and Townsend, 2003; Croll et al., 2005; Chae et al., 2008; Suntsov and Brodeur, 2008; Hipfner, 2009). If N. nomurai blooms contribute to a reduction in the krill population, the food supply of higher trophic-level animals may also decrease. In other words, N. nomurai blooms may affect krill abundance as a result of competition for food resources, and this impact may be transmitted to higher trophic-level organisms.
The stable isotope bi-plot showed that N. nomurai shared similar δ13C and δ15N values with krill and small copepods (Fig. 4), which indicated that the effect of N. nomurai blooms on krill and small copepods was likely caused mainly by competition for food resources. However, the small copepod O. similis may also be preyed upon by N. nomurai, which indicates overlap between the δ15N values of N. nomurai and its prey. This phenomenon has been reported in P. noctiluca and A. aurita (D'Ambra et al., 2013; Milisenda et al., 2018). Therefore, N. nomurai likely influences small copepods through predation and competition. Because arrow worms and fish larvae are also predators of copepods, they could be indirectly affected by N. nomurai. In summary, the impact of N. nomurai blooms on zooplankton communities exerts a negative bottom-up influence that is mainly a result of diminishing the low trophic-level prey.
4.3 Limitations and countermeasures of SIAR tools used in this studyTwo issues must be considered when using SIAR. First, to obtain accurate results, the source data should include as many of the consumer's total food sources as possible (Remon et al., 2016). Yet if all of N. nomurai possible food sources were substituted into the model, the number of food sources would be too large, which can influence the model's accuracy (Parnell et al., 2010). Secondly, the contribution of two relatively close food sources with different abundances yet similar isotope values to the diet of consumers should be different. However, because the data imported to the SIAR model includes only isotope values that do not consider abundance (Parnell et al., 2010), similar isotope values substituted into the model may result in similar contribution ratios; this is not consistent with real-world observations. To solve these two problems, we chose to group sources according to particle size. After investigating zooplankton abundance in the study area, the isotopic data of dominant species in each group were selected to execute the SIAR model. The number of food sources was reduced as few as possible to ensure successful operation of the model. The food sources data can also represent the vast majority of the food for N. nomurai, ensuring the accuracy of results. Future research efforts shall focus on increasing the sampling frequency to obtain stable isotopic data for more species and establishing a stable isotope database for plankton. After that, plankton could be classified into groups according to the stable isotope data to decrease the number of sources imported into the model. When running SIAR models, substituting each group of plankton as a single source into the model; that way, the errors caused by different species abundances can be avoided. In future, this database may also facilitate the study of material energy flows in marine research.
5 CONCLUSIONFood compositions of large and small N. nomurai are similar. Both are mostly composed by organisms with size smaller than 1 000 μm in diameter. Therefore, the feeding pressure of N. nomurai blooms is mostly put on small planktons. The impacts on large zooplanktons (e.g. krills) are mainly caused by food competition. This would provide certain guiding significance for the impact assessment research of N. nomurai bloom. In addition, the food composition of N. nomurai could be reference materials for the forecasting research of N. nomurai blooms.
6 DATA AVAILABILITY STATEMENTThe data that support the findings of the current study are available on reasonable request from the corresponding author.
7 ACKNOWLEDGMENTWe thank Drs. Song FENG, Lijuan WANG, Pengpeng WANG, and Nan YU, from Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, for their great assistance during shipboard research activities.
Allan E L, Ambrose S T, Richoux N B, Froneman P W. 2010. Determining spatial changes in the diet of nearshore suspension-feeders along the South African coastline: stable isotope and fatty acid signatures. Estuarine, Coastal and Shelf Science, 87(3): 463-471.
DOI:10.1016/j.ecss.2010.02.004 |
Bargu S, Lefebvre K, Silver M W. 2006. Effect of dissolved domoic acid on the grazing rate of krill Euphausia pacifica. Marine Ecology Progress Series, 312: 169-175.
DOI:10.3354/meps312169 |
Bray J R, Curtis J T. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecology Monography 27. p. 326-349.
|
Brinton E, TownsendA. 2003. Decadal variability in abundances of the dominant euphausiid species in southern sectors of the California Current. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 50(14-16): 2449-2472.
DOI:10.1016/S0967-0645(03)00126-7 |
Budge S M, Parrish C C. 1998. Lipid biogeochemistry of plankton, settling matter and sediments in Trinity Bay, Newfoundland. Ⅱ. Fatty acids. Organic Geochemistry, 29(5-7): 1547-1559.
DOI:10.1016/S0146-6380(98)00177-6 |
Cabana G, Rasmussen J B. 1996. Comparison of aquatic food chains using nitrogen isotopes. Proceedings of the National Academy of Sciences of the United States of America, 93(20): 10844-10847.
DOI:10.1073/pnas.93.20.10844 |
Chae J, Han C, Lee J H, Hong J S. 2008. A short note on a euphausiid, Euphausia pacifica, an important food source to demersal fishes in Uljin area, the eastern coast of Korea. Journal of Environmental Biology, 29(4): 471-474.
|
Connelly T L, Deibel D, Parrish C C. 2014. Trophic interactions in the benthic boundary layer of the Beaufort Sea shelf, Arctic Ocean: combining bulk stable isotope and fatty acid signatures. Progress in Oceanography, 120: 79-92.
DOI:10.1016/j.pocean.2013.07.032 |
Croll D A, Marinovic B, Benson S, Chavez F P, Black N, Ternullo R, Tershy B R. 2005. From wind to whales: trophic links in a coastal upwelling system. Marine Ecology Progress Series, 289: 117-130.
DOI:10.3354/meps289117 |
Cui Y, Wu Y, Zhang J, Wang N. 2012. Potential dietary influence on the stable isotopes and fatty acid compositions of jellyfishes in the Yellow Sea. Journal of the Marine Biological Association of the United Kingdom, 92(6): 1325-1333.
DOI:10.1017/S0025315412000082 |
D'Ambra I, Graham W M, Carmichael R H, Malej A, Onofri V. 2013. Predation patterns and prey quality of medusae in a semi-enclosed marine lake: implications for food web energy transfer in coastal marine ecosystems. Journal of Plankton Research, 35(6): 1305-1312.
DOI:10.1093/plankt/fbt065 |
Dalsgaard J, St John M, Kattner G, Müller-Navarra D, Hagen W. 2003. Fatty acid trophic markers in the pelagic marine environment. Advances in Marine Biology, 46: 225-340.
DOI:10.1016/S0065-2881(03)46005-7 |
DeNiro M J, Epstein S. 1978. Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimica Acta, 42(5): 495-506.
DOI:10.1016/0016-7037(78)90199-0 |
Dilling L, Wilson J, Steinberg D, Alldredge A. 1998. Feeding by the euphausiid Euphausia pacifica and the copepod Calanus pacificus on marine snow. Marine Ecology Progress Series, 170: 189-201.
DOI:10.3354/meps170189 |
Ding FY, Cheng J H. 2005. The analysis on fish stock characteristics in the distribution areas of large jellyfish during summer and autumn in the East China Sea region. Marine Fisheries, 27(2): 120-128.
(in Chinese with English abstract) |
El-Sabaawi R, Dower J F, Kainz M, Mazumder A. 2009. Characterizing dietary variability and trophic positions of coastal calanoid copepods: insight from stable isotopes and fatty acids. Marine Biology, 156(3): 225-237.
DOI:10.1007/s00227-008-1073-1 |
Folch J, Lees M, Stanley S G H. 1957. A simple method for the isolation and purification of total lipides from animal tissues. Journal of Biological Chemistry, 226(1): 497-509.
DOI:10.1016/S0021-9258(18)64849-5 |
Gorsky G, Ohman M D, Picheral M, Gasparini S, Stemmann L, Romagnan J B, Cawood A, Pesant S, García-Comas C, Prejger F. 2010. Digital zooplankton image analysis using the ZooScan integrated system. Journal of Plankton Research, 32(3): 285-303.
DOI:10.1093/plankt/fbp124 |
Grosjean P, Picheral M, Warembourg C, Gorsky G. 2004. Enumeration, measurement, and identification of net zooplankton samples using the ZOOSCAN digital imaging system. ICES Journal of Marine Science, 61(4): 518-525.
DOI:10.1016/j.icesjms.2004.03.012 |
Gueroun S K M, Molinero J C, Piraino S, Yahia M N D. 2020. Population dynamics and predatory impact of the alien jellyfish Aurelia solida (Cnidaria, Scyphozoa) in the Bizerte Lagoon (southwestern Mediterranean Sea). Mediterranean Marine Science, 21(1): 22-35.
DOI:10.12681/mms.17358 |
Henschke N, Everett J D, Suthers I M, Smith J A, Hunt B P V, Doblin M A, Taylor M D. 2015. Zooplankton trophic niches respond to different water types of the western Tasman Sea: a stable isotope analysis. Deep Sea Research Part Ⅰ: Oceanographic Research Papers, 104: 1-8.
DOI:10.1016/j.dsr.2015.06.010 |
Hipfner J M. 2009. Euphausiids in the diet of a North Pacific seabird: annual and seasonal variation and the role of ocean climate. Marine Ecology Progress Series, 390: 277-289.
DOI:10.3354/meps08209 |
Hu W Y, Chen J, Li D Y, Wang A J, Li W A, Huang C B. 2017. Composition and particle size change of suspended particulate matter in spring and autumn in the northwest Pacific Ocean. Journal of Applied Oceanography, 36(2): 221-232.
(in Chinese with English abstract) |
Ishii H, Tanaka F. 2001. Food and feeding of Aurelia aurita in Tokyo Bay with an analysis of stomach contents and a measurement of digestion times. Hydrobiologia, 451(1-3): 311-320.
DOI:10.1023/A:1011814525325 |
Kaneda T. 1991. Iso- and anteiso-fatty acids in bacteria: biosynthesis, function, and taxonomic significance. Microbiological Reviews, 55(2): 288-302.
DOI:10.1128/mr.55.2.288-302.1991 |
Kattner G, Fricke H S G. 1986. Simple gas-liquid chromatographic method for the simultaneous determination of fatty acids and alcohols in wax esters of marine organisms. Journal of Chromatography A, 361: 263-268.
DOI:10.1016/S0021-9673(01)86914-4 |
Kawahara M, Uye S I, Ohtsu K, Iizumi H. 2006. Unusual population explosion of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) in East Asian waters. Marine Ecology Progress Series, 307: 161-173.
DOI:10.3354/meps307161 |
Kling G W, Fry B, O'Brien W J. 1992. Stable isotopes and planktonic trophic structure in Arctic lakes. Ecology, 73(2): 561-566.
DOI:10.2307/1940762 |
Lee H E, Yoon W D, Lim D. 2008. Description of feeding apparatus and mechanism in Nemopilema nomurai Kishinouye (Scyphozoa: Rhizostomeae). Ocean Science Journal, 43(1): 61-65.
DOI:10.1007/BF03022432 |
Li C L, Yang G, Ning J, Sun J, Yang B, Sun S. 2013. Response of copepod grazing and reproduction to different taxa of spring bloom phytoplankton in the Southern Yellow Sea. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 97: 101-108.
DOI:10.1016/j.dsr2.2013.05.018 |
Liu X J, Yu Z M, Song X X, Cao X H. 2010. Distribution in the stable nitrogen isotope of the suspended particulate organic matter in the Yangtze River (Changjiang) estuary, China and its biogeochemistry implications. Marine Sciences, 34(1): 11-17.
(in Chinese with English abstract) |
Lo W T, Chen I L. 2008. Population succession and feeding of Scyphomedusae, Aurelia aurita, in a eutrophic tropical lagoon in Taiwan. Estuarine, Coastal and Shelf Science, 76(2): 227-238.
DOI:10.1016/j.ecss.2007.07.015 |
Logan J M, Jardine T D, Miller T J, Bunn S E, Cunjak R A, Lutcavage M E. 2008. Lipid corrections in carbon and nitrogen stable isotope analyses: comparison of chemical extraction and modelling methods. Journal of Animal Ecology, 77(4): 838-846.
DOI:10.1111/j.1365-2656.2008.01394.x |
Milisenda G, Rossi S, Vizzini S, Fuentes V L, Purcell J E, Tilves U, Piraino S. 2018. Seasonal variability of diet and trophic level of the gelatinous predator Pelagia noctiluca (Scyphozoa). Scientific Reports, 8(1): 12140.
DOI:10.1038/s41598-018-30474-x |
Nakagawa Y, Ota T, Endo Y, Taki K, Sugisaki H. 2004. Importance of ciliates as prey of the euphausiid Euphausia pacifica in the NW North Pacific. Marine Ecology Progress Series, 271: 261-266.
DOI:10.3354/meps271261 |
Odum E P. 1959. Fundamentals of Ecology. Saunders, Philadelphia. 384p.
|
Ohman M D. 1984. Omnivory by Euphausia pacifica: the role of copepod prey. Marine Ecology Progress Series, 19: 125-131.
DOI:10.3354/meps019125 |
Omori M, Kitamura M. 2004. Taxonomic review of three Japanese species of edible jellyfish (Scyphozoa: Rhizostomeae). Plankton Biology & Ecology, 51(1): 36-51.
|
Parnell A C, Inger R, Bearhop S, Jackson A L. 2010. Source partitioning using stable isotopes: coping with too much variation. PLoS One, 5(3): e9672.
DOI:10.1371/journal.pone.0009672 |
Parrish C C, Mckenzie C H, Macdonald B A, Hatfield E A. 1995. Seasonal studies of seston lipids in relation to microplankton species composition and scallop growth in South Broad Cove, Newfoundland. Marine Ecology Progress Series, 129: 151-164.
DOI:10.3354/meps129151ISBN:0171-8630 |
Pitt K A, Connolly R M, Meziane T. 2009. Stable isotope and fatty acid tracers in energy and nutrient studies of jellyfish: a review. Hydrobiologia, 616(1): 119-132.
DOI:10.1007/s10750-008-9581-z |
Post D M. 2002. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology, 83(3): 703-718.
DOI:10.1890/0012-9658(2002)083[0703:USITET]2.0.CO;2 |
Qin Y S, Li F, Xu S M, John M, Richard L. 1988. Study on suspended matter in seawater in the Southern Yellow Sea. Chinese Journal of Oceanology and Limnology, 6(3): 201-2015.
DOI:10.1007/BF02846498 |
Remon J, Bower D S, Gaston T F, Clulow J, Mahony M J. 2016. Stable isotope analyses reveal predation on amphibians by a globally invasive fish (Gambusia holbrooki). Aquatic Conservation: Marine and Freshwater Ecosystems, 26(4): 724-735.
DOI:10.1002/aqc.2631 |
Schukat A, Auel H, Teuber L, Lahajnar N, Hagen W. 2014. Complex trophic interactions of calanoid copepods in the Benguela upwelling system. Journal of Sea Research, 85: 186-196.
DOI:10.1016/j.seares.2013.04.018 |
Sun S, Huo Y Z, Yang B. 2010. Zooplankton functional groups on the continental shelf of the yellow sea. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 57(11-12): 1006-1016.
DOI:10.1016/j.dsr2.2010.02.002 |
Sun S, Zhang F, Li C L, Wang S W, Wang M X, Tao Z C, Wang Y T, Zhang G T, Sun X X. 2015. Breeding places, population dynamics, and distribution of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) in the Yellow Sea and the East China Sea. Hydrobiologia, 754(1): 59-74.
DOI:10.1007/s10750-015-2266-5 |
Suntsov A V, Brodeur R D. 2008. Trophic ecology of three dominant myctophid species in the northern California Current region. Marine Ecology Progress Series, 373: 81-96.
DOI:10.3354/meps07678 |
Tang Q, Jin X, Zheng Y, Meng T. 2006. Marine living resources and inhabited environment of Chinese exclusive economic zones. Science Press, Beijing. 3-28.
(in Chinese)
|
Teuber L, Schukat A, Hagen W, Auel H. 2014. Trophic interactions and life strategies of epi- to bathypelagic calanoid copepods in the tropical Atlantic Ocean. Journal of Plankton Research, 36(4): 1109-1123.
DOI:10.1093/plankt/fbu030 |
Tilves U, Fuentes V L, Milisenda G, Parrish C C, Vizzini S, Sabatés A. 2018. Trophic interactions of the jellyfish Pelagia noctiluca in the NW Mediterranean: evidence from stable isotope signatures and fatty acid composition. Marine Ecology Progress Series, 591: 101-116.
DOI:10.3354/meps12332 |
Tilves U, Purcell J E, Fuentes V L, Torrents A, Pascual M, Raya V, Gili J M, Sabatés A. 2016. Natural diet and predation impacts of Pelagia noctiluca on fish eggs and larvae in the NW Mediterranean. Journal of Plankton Research, 38(5): 1243-1254.
DOI:10.1093/plankt/fbw059 |
Uye S I. 2008. Blooms of the giant jellyfish Nemopilema nomurai: a threat to the fisheries sustainability of the East Asian Marginal Seas. Plankton and Benthos Research, 3(S1): 125-131.
DOI:10.3800/pbr.3.125 |
Uye S I. 2011. Human forcing of the copepod-fish-jellyfish triangular trophic relationship. Hydrobiologia, 666(1): 71-83.
DOI:10.1007/s10750-010-0208-9 |
Uye S I. 2014. The Giant Jellyfish Nemopilema nomurai in East Asian Marginal Seas. In: Pitt K A, Lucas C H eds. Jellyfish Blooms. Springer, Dordrecht. p. 185-205, https://doi.org/10.1007/978-94-007-7015-7_8.
|
Wang S W, Budge S M, Iken K, Gradinger R R, Springer A M, Wooller M J. 2015. Importance of sympagic production to Bering Sea zooplankton as revealed from fatty acid-carbon stable isotope analyses. Marine Ecology Progress Series, 518: 31-50.
DOI:10.3354/meps11076 |
Wu Y, Li H Y, Li S F, Cheng J H. 2008. Review on the current situation of macro-jellyfish research and expectations. Marine Fisheries, 30(1): 80-87.
(in Chinese with English abstract) |
Yang G, Li C L, Guilini K, Peng Q C, Wang Y Q, Zhang Y, Zhang Y S. 2016. Feeding strategies of four dominant copepod species in Prydz Bay, Antarctica: insights from a combined fatty acid biomarker and stable isotopic approach. Deep Sea Research Part I: Oceanographic Research Papers, 114: 55-63.
DOI:10.1016/j.dsr.2016.04.016 |
Yoon W D, Lee H E, Han C, Chang S J, Lee K. 2014. Abundance and distribution of Nemopilema nomurai (Scyphozoa, Rhizostomeae), in Korean waters in 2005-2013. Ocean Science Journal, 49(3): 183-192.
DOI:10.1007/s12601-014-0018-5 |
Zhang F, Sun S, Jin X S, Li C L. 2012. Associations of large jellyfish distributions with temperature and salinity in the Yellow Sea and East China Sea. Hydrobiologia, 690(1): 81-96.
DOI:10.1007/s10750-012-1057-5 |