 2021, Vol. 39
2021, Vol. 39Institute of Oceanology, Chinese Academy of Sciences
Article Information
- TIAN Yuan, HU Simin, LIN Xianzhi, HUANG Hui, SONG Xingyu, YAN Yan, XIE Xuedong, LI Tao, LIU Sheng
- Mechanisms of high-frequency dinoflagellate blooms of Scrippsiella trochoidea in Daya Bay, South China Sea
- Journal of Oceanology and Limnology, 39(4): 1293-1304
- http://dx.doi.org/10.1007/s00343-020-9082-0
Article History
- Received Mar. 27, 2019
- accepted in principle Sep. 22, 2019
- accepted for publication Aug. 6, 2020




2 University of Chinese Academy of Sciences, Beijing 100049, China;
3 Tropical Marine Biological Research Station in Hainan, Chinese Academy of Sciences, Sanya 572000, China;
4 Nansha Marine Ecological and Environmental Research Station, Chinese Academy of Sciences, Guangzhou 510301, China;
5 Guangdong Ocean and Fishery Environment Monitoring and Forecasting Center, Guangzhou 510222, China
Harmful algal blooms (HABs) have adverse impacts on the aquatic environment, damage the stabilization of the ecosystem, and even give rise to severe economic losses around the world (Glibert et al., 2018). About 300 species of phytoplankton have ever caused "red tides", yet only 60–80 species (20%–26.67%) of them are actually harmful or toxic, in which dinoflagellates (45–60 taxa) accounts for about 75% of all HAB species (Smayda, 1997). Algae blooms could affect the water quality, weaken light penetration and reduce the dissolved oxygen rapidly to form low-oxygen "dead-zones" during bloom degradation. As a consequence, benthic organisms would be negatively influenced (Wells et al., 2015). More seriously, dinoflagellate blooms may influence the ecosystem structure with copepods decreasing and small jellyfish increasing, which will lead to the abnormal transmission process of the food chain and eventually disrupt the stability of the whole ecosystem (Lin et al., 2014). Moreover, some dinoflagellate species are the producers of toxins and such blooms can be lethal to aquatic organisms, causing significant economic losses to the aquaculture (Shin et al., 2017; Townhill et al., 2018) and even affecting human health (Backer and McGillicuddy, 2006).
Dinoflagellate blooms along the coastal waters of China presented some regional characteristics. In Bohai Sea, the dinoflagellate blooms species were diverse, including Noctiluca scintillans, Ceratium spp. and Gymnodinium spp. In general, N. scintillans blooms frequently occurred in the west of Liaodong Bay and Qinhuangdao coastal area. While Ceratium spp. and Gymnodinium spp. bloomed frequently in Bohai Bay (Lin et al., 2017). In the East China Sea, Prorocentrum donghaiense periodically bloomed over an extremely large area (~10 000 km2) after 2000 (Lu and Goebel, 2001). In the South China Sea, the dominant bloom species were N. scintillans, Scrippsiella trochoidea, and Karenia mikimotoi, occupying about 34.53%, 16.14%, and 6.28% of total dinoflagellate blooms, respectively (Li et al., 2019; Tian et al., 2020). Dinoflagellate blooms showed diverse patterns in different regions (Glibert et al., 2018). It is essential to understand the characteristics and mechanisms of the outbreak of dinoflagellate blooms for effective environmental management and socio-economic development.
Scrippsiella trochoidea are widely distributed all over the world, especially in the tropical and subtropical regions between 40°N and 40°S, and the higher cell density were usually occurred in the eutrophic waters in coastal area (Hallegraeff, 1992) and usually bloomed in shallow waters (about 10-m depth) in the inner bay (Mulholland et al., 2018). It could survive at the temperature of 10–35.2 ℃ and the salinity of 2.3–55 (Kim and Han, 2000; Hameed and Saburova, 2015). The optimum growth temperature of S. trochoidea varies with regions. It had been reported as 26.5–27 ℃ in the southwestern Gulf of California (Gárate-Lizárraga et al., 2009) and 25 ℃ in Yongil Bay in South Korea (Kim and Han, 2000), which was field investigation. The laboratory temperature experiment showed 22 ℃ and 25 ℃ for S. trochoidea strain isolated from North Yellow Sea (Xu and Pang, 2012) and Zhoushan Sea (Xu et al., 2004). Usually, its bloom occurred in 25–30 ℃ (Mulholland et al., 2018). Moreover, S. trochoidea could produce mass cysts under adverse environmental conditions, forming a massive source for its bloom (Xiao et al., 2001). However, the cysts of S. trochoidea were widespread but the S. trochoidea blooms occurred in a certain area only (Kim and Han, 2000). Recent studies show that, the causes of S. trochoidea blooms were diverse for nutrient supply in different regions, such as the upwelling (nutrient input) in southern Gulf of Mexico (Licea et al., 2000) and the total nitrogen increasing in Kochi estuary of southeastern Arabian Sea (Kumar et al., 2018). However, the initiation and persistence of S. trochoidea blooms in Port Shelter, Hong Kong, China were not directly driven by the changes in nutrients (Yin et al., 2008). In addition, S. trochoidea blooms might be triggered by the combination of strong light and high iron concentration based on laboratory experiments (Cai et al., 2009).
Daya Bay is a semi-closed subtropical bay with frequent occurrence of HAB in the northern South China Sea. Previous studies showed that the increasing anthropogenic disturbances (e.g. coastal aquaculture industry) might be the most crucial trigger for high frequency HAB (Yu et al., 2007). First, the inner bays (such as Yaling Bay and Dapeng Cove) were in the mesotrophic status and tended to eutrophication (Huan et al., 2016). Secondly, the Daya Bay Nuclear Power Plant (DNPP) and Ling-ao Nuclear Power Plant (LNPP) discharged a mass of thermal effluent to adjacent waters (about 285 m3/s) and affected as far as 8–10 km away from south of the nuclear power plant (Tang et al., 2003; Hao et al., 2016). In the affected waters, the water temperature significantly increased, and the phytoplankton community structure had shown a tendency to change from diatom-dominate to dinoflagellate-dominate (Li et al., 2011). Moreover, an increasing dinoflagellate blooms, especially S. trochoidea blooms had been frequently reported in the inner bay of Daya Bay since 1998, especially in Yaling Bay and Dapeng Cove (Tian et al., 2020). S. trochoidea produced a great deal of resting cysts and the cysts could germinate at appropriate status, which could be the species source for the long-term blooms (Xiao et al., 2003; Yu et al., 2007). However, it is still unclear how these factors coordinately act during the formation of dinoflagellate blooms. In this study, data from historical HABs, field investigation, and laboratory experiments were combined to explore the potential process of S. trochoidea blooms in Daya Bay.
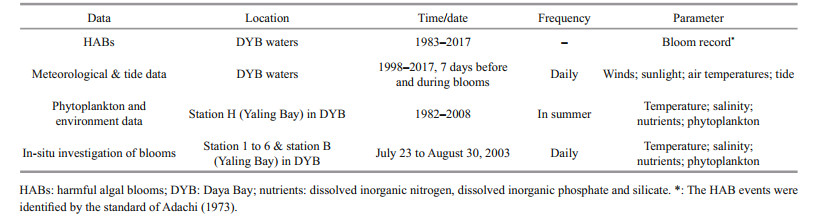
2 MATERIAL AND METHOD 2.1 Data collectionTo explore the variation trends of occurrence patterns of dinoflagellate HABs and phytoplankton community, HAB records (1983–2017), long-term environmental data (1982–2008), and possible influence parameters were collected (Table 1). HAB records were collected from the Guangdong Ocean and Fishery Environment Monitoring and Forecasting Center, Guangdong Provincial Marine Environmental Quality Bulletin and previously published literatures (Qian et al., 2000; Lin and Liang, 2002). Meteorological factors were gathered from the Milos 520 automatic meteorological station (37.3 m above sea level) located at 22°31′N, 114°31′E (Fig. 1a). Tide data were taken from the tide table prepared by the China National Ocean Information Center. Phytoplankton and environmental data in long-term series were obtained from Marine Biology Research Station of Daya Bay, the South China Sea Institute of Oceanology, Chinese Academy of Sciences.

|
| Fig.1 Location of sampling stations DNPP: Daya Bay Nuclear Power Plant; LNPP: Ling-ao Nuclear Power Plant. |
To study the factors influencing the process of dinoflagellate blooms, we also tracked a S. trochoidea bloom in Yaling Bay from August 13 to 30, 2003. Before bloom, from July 23 to August 13, data were collected at 6 stations (station 1 to 6) every 5 days (Fig. 1b). A mixed bloom of S. trochoidea and Chattonella marina was first observed at station B on August 13. From August 14, the investigation was performed daily exclusive of the 22, 24, and 25 due to a typhoon (Table 1).
500-mL samples were collected from surface water and fixed in Lugol's solution for phytoplankton observation. 50-mL aliquots were then concentrated to 1 mL and phytoplankton species were identified and counted in a Sedgwick-Rafter counting chamber using an Olympus BX51 microscope. 20–50 columns were randomly counted from 200 columns in Sedgwick-Rafter counting chamber, and the abundance of the algae in the original water sample was obtained according to the concentration ratio (Lin et al., 2012). Each sample was identified to species level whenever possible. For nutrient analysis, 50-mL samples were filtered through 0.45-μm cellulose acetate membrane filters, and the filters were then placed in polyethylene plastic bottles and immediately frozen at -20 ℃. A Skalar auto-analyzer (Skalar Analytical B.V. SanPlus, Netherlands) was used to measure nutrient concentrations in the laboratory (Knap et al., 1996). The dissolved inorganic nitrogen (DIN) content was calculated based on the combined total nitrate (NO3-), nitrite (NO2-) and ammonium (NH4+), while the dissolved inorganic phosphorus (DIP) content was measured as phosphate (PO43-). Water temperature, salinity, pH and water depth were measured on-site using a multi-parameter water quality monitor (YSI 6600V2, USA).
2.3 Temperature adaptation of S. trochoideaScrippsiella trochoidea was obtained from the algae species library of Jinan University, which was initially isolated from Dapeng Bay, South China Sea. In order to verify the temperature adaptation of S. trochoidea, four temperature treatments with replicate were performed at 27±1, 30±1, 33±1, and 36±1 ℃. All treatments were maintained in a zone light incubator with L1 medium (FPG3, Ningbo Laifu Technology Limited Company, China). The light intensity was 100 μmol/(m2·s) with 12 h꞉12 h light꞉ dark cycle. One-milliliter samples were taken daily and fixed in Lugol's solution, then placed in a Sedgwick-Rafter counting chamber and counted under an Olympus BX51 microscope as described in Section 2.2.
2.4 Data treatment and analysisMixed blooms formed by two or more species were regarded as multiple blooms occurring concurrently in this study. The available potential energy (APE, J/m3) had previously been used to study the mixing processes in stratified fluids and used as an indicator of water stratification (Kang and Fringer, 2010). APE was established using the equations below:
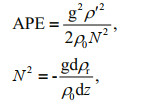
where ρ' is the perturbation density; ρ0 is the constant reference density associated with the Boussinesq approximation; ρr the reference density; z the depth of water column; and N the buoyancy frequency.
Since phytoplankton sampling was a long-term process, involving the classification of phytoplankton by routine microscopy, even for experienced taxonomists, identification to the species level may not be reliable (Godhe et al., 2015). Therefore, we employed the genus level of phytoplankton, studied the relationship between the phytoplankton and environmental parameters (temperature, pH, salinity, APE, DIN, DIP and silicate) with canonical correlation analysis (CCA), and determined the relationship between the abundance of S. trochoidea and environmental parameters during the bloom period using the Pearson correlation coefficient. The algae growth rate (μ) was resolved by the equation (Sun and Ning, 2005):
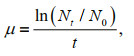
where, N0 is the initial cell density, and Nt is the cell density after t time.
Statistical analysis was executed with R framework 3.4.1 (R Core Team 2016) and mapping was performed using SigmaPlot 12.5 and Golden Software Surfer 13.0.
3 RESULT 3.1 Prevalence of dinoflagellate blooms in Daya BayIn total, 94 HABs, caused by approximately 36 species, were observed in Daya Bay from 1983 to 2017. If mixed HABs were counted as individual events occurring concurrently in the same place, 111 blooms were identified. Generally, the frequency of HABs increased from less than 5/a in the 1980s to 10/a in 1991, reaching peak in 2000 (17/a). After 2010, although the HABs decreased by 65%–94% compared to 2000, the frequency was 3.1/a on average. Remarkably, the percentage of dinoflagellate blooms increased in a stepwise manner with the operation of DNPP, Phase I and Phase II of LNPP in 1994, 2003, and 2011 (Fig. 2b), from 15% before 1994 to 35% between 1994–2002, then to 50% between 2003 and 2010, and to 77% between 2011 and 2017. The percentage of diatom blooms decreased from 60% before 1994 to 45% between 1994 and 2002, and 5% after 2003. Numerically, before 2003, the diatom blooms and dinoflagellate blooms were in the same order with 35 and 21 times, respectively. However, after 2003, the diatom blooms almost disappeared (twice) and the dinoflagellate blooms became the most crucial species of HABs (26 times) (Fig. 2a).

|
| Fig.2 Frequency of HABs in Daya Bay from 1983 to 2017 a. outbreak status of different HABs; b. occurrence probability of different HABs. The Daya Bay Nuclear Power Plant (began operation in 1994; the first-stage and second-stage of the Ling-ao Nuclear Power Plant began to operate in 2003 and 2011, respectively. NPPU: nuclear power plant unit; TD: thermal discharge. |
Interestingly, dinoflagellate blooms, especially S. trochoidea blooms, appeared most frequently in summer. First, the HAB-prone area was the western part of Daya Bay, with 59 cases (53%) occurring in Yaling Bay and 8 cases (7%) in Dapeng Cove. Secondly, in summer, dinoflagellate blooms (19 cases) accounted for 58% of HABs in this area, including 16 S. trochoidea bloom events. In Yaling Bay, the HAB prone area, dinoflagellate blooms accounted for 77% of the total blooms in summer, while S. trochoidea blooms accounted for 90% of all dinoflagellate blooms.
Scrippsiella trochoidea was the main dinoflagellate bloom forming species (25 in total), which occurred almost every year since the first observation in Daya Bay in 1998. Moreover, the chances of S. trochoidea blooms also increased after the operation of DNPP. Before 2003, S. trochoidea bloomed 9 times, accounting for 13% and 43% of all blooms and all dinoflagellate blooms, respectively. Between 2003 and 2010, S. trochoidea blooms events were 8, accounted for 44% and 89% of the total and dinoflagellate blooms, respectively. After 2010, the number was 8. Although the proportion of S. trochoidea blooms decreased, they still accounted for 36% of the total blooms and 47% of dinoflagellate blooms. S. trochoidea gradually became the main species of dinoflagellate blooms in Daya Bay in the past decades (Fig. 2b).
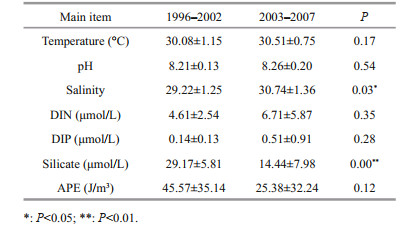
3.2 Environmental change in the bloom-prone areaYaling Bay is a shallow inner bay (~6 m), and is easily affected by tide and wind. Taking 2003 as the watershed for bloom change, the environment change in the bloom-prone area in summer was compared (Table 2). The water temperature increased slightly and stayed above 30 ℃, and there was no significant difference between the two periods (P > 0.05). Both DIN and DIP increased after 2003, although they were not significantly different to those before 2003 (P > 0.05). Remarkably, the silicate concentrations in Yaling Bay significantly reduced (P < 0.001) after 2003, from 29.17±5.81 to 14.44±7.98 μmol/L. The salinity significantly increased (P < 0.05) after 2003, from 29.22±1.25 to 30.74±1.36. The APE decreased from 45.57±35.14 to 25.38±32.24 J/m3, and had no significant difference between the two periods (P > 0.05).
CCA analysis was performed for the associations between phytoplankton genus abundance and environment factors during summer, in the dinoflagellate bloom-prone area (Yaling Bay). 75 data sets collected from 1996 to 2007 were analyzed (Fig. 3). Diatoms (Bacillariophyta) were widely dispersed and were not affected by specific environmental factors. Cyanobacteria and Raphidophyceae were distributed more frequently in the areas where the temperature varies. Dinoflagellates (Pyrrophyta) were mostly found near the temperature axis and positively increased with temperature. Specifically, the genus Scrippsiella was located near the temperature axis and the trend of its abundance was consistent with dinoflagellates. Only one species (S. trochoidea) belonging to the genus Scrippsiella was observed in Daya Bay. A consistent trend was observed between changes in temperature and the relative abundance of dinoflagellates (R2=0.37, P=0.04) (Fig. 3b). Especially in July 1998, August 1999, August 2001, and August 2004, high temperature and high dinoflagellates abundance were observed at the same time. In August 2004, when the water temperature was as high as 32.4 ℃, the relative abundance of dinoflagellates reached the maximum of 55%.

|
| Fig.3 The relationship between phytoplankton and environmental factors in Yaling Bay from 1996 to 2007 a. canonical correlation analysis (CCA) of phytoplankton genus abundances (response factors) and environmental factors (predictor factors); b. linear regression of percentage of dinoflagellate cell abundance and temperature. Scaling of the CCA biplot was set to "2" to retain the correct angles (correlations) between factors. Si: silicate; T: temperature; S: salinity. Squares represented Bacillariophyta; circles represented Pyrrophyta; triangles represented Chrysophyta; rhombus represented Cyanophyta; inverted triangles represented Raphidophyceae. |
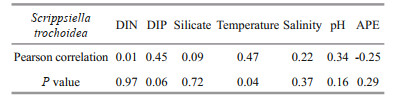
A mixed bloom of S. trochoidea and C. marina occurred in Yaling Bay on August 13–30, 2003. Correlation analysis showed that the abundance of S. trochoidea was only significantly correlated with temperature (P < 0.05), with a Pearson correlation coefficient of 0.47, and not correlated with other environmental factors (DIN, DIP, silicate, salinity, pH, and APE) (P > 0.05) (Table 3). The water temperature fluctuated between 28.2 and 32.1 ℃ during the bloom period and the maximum cell density of S. trochoidea reached 5.59×107 cells/L at 32.1 ℃.
|
In laboratory incubation, S. trochoidea could grow normally at 27, 30, and 33 ℃, but could not survive at 36 ℃. The average growth rates at 27, 30, and 33 ℃ were 0.32±0.02, 0.34±0.04, and 0.33±0.05/d, respectively. S. trochoidea grew better at 30 ℃ than at other temperatures and quickly reached the exponential growth phase (on the 6th day), with maximum cell density (1.01±0.10)×105 cells/mL on the 10th day.
3.5 Tide and weather conditions during S. trochoidea bloomsThe sunlight hours increased significantly before the S. trochoidea blooms, which was 3.86±0.86, 4.47±0.85, 7.37±0.76, 8.61±0.77, and 9.43±0.35 h/d on days 4, 3, 2, and 1 before blooms and on the first day of blooms, respectively. The air temperatures also increased to 27.62±0.35, 27.93±0.38, 28.46±0.42, 28.58±0.39, and 28.71±0.36 ℃ on days 4, 3, 2, and 1 before bloom and on the first day of bloom, respectively.
Of the 20 defined blooms, 14 blooms (70%) occurred during the monthly spring tide period and/or periods of strong winds (daily average wind velocity ˃7 m/s) (Supplementary Table S1). After spring tide period and/or periods of high wind, 65% of the daily average wind velocity was less than 3.3 m/s, similarly to 74% of the daily average wind velocity during the bloom period. Both 7 days before the onset of blooms and during bloom periods, the wind directions were mostly south-southwest (SSW).
4 DISCUSSIONBecause of the ongoing global warming, the seawater temperature has increased by 0.5 ℃ over the past 100 year (IPCC, 2007). Higher temperature was beneficial for dinoflagellate by intensifying the water stratification and enhancing stabilization of water column (Jones and Gowen, 1990; Hinder et al., 2012). Dinoflagellate could grow better in stable waters. Their complicated trophic ways (e.g. photoautotrophy, heterotrophy and mixotrophy) enhanced their competitive advantage over diatoms (Burkholder and Glasgow, 1997). Numerous field studies had suggested that dinoflagellate dominancy might be one of main reasons for more and more dinoflagellate blooms globally (Li et al., 2011; Godhe et al., 2015; Xiao et al., 2018). Furthermore, the life cycle of dinoflagellate was complicated. They can generate resting cysts under unfavorable environment conditions, which can be kept in sediment for a long time and germinate in short time when environment is suitable. This could be a seed resource for dinoflagellate blooms in appropriate environment, such as Akashiwo Sanguinea (Tang and Gobler, 2015). Therefore, the frequency, duration, and intensity of dinoflagellate blooms are related to biological and physical-chemical factors. The causes and pattern of HABs, especially dinoflagellate blooms, are extremely complicated. Even a multitude of studies were conducted, it was still difficult to predict accurately (Smayda and Reynolds, 2003). Taking into consideration the environmental parameters and atmospheric conditions, we proposed a potential mechanism of S. trochoidea blooms in this study, and provided an insight for understanding the high frequency of dinoflagellate blooms in coastal areas.
4.1 Environment change benefit dinoflagellate growthDue to the rapid socio-economic development, the coastal waters have been seriously affected by human activities. Daya Bay is a semi-closed bay, and gradually shifted from oligotrophic level before the 1990s to mesotrophic level after 2000. Some inner bays (Yaling Bay, Dapeng Cove, etc.) showed a trend of eutrophication (Huan et al., 2016). Moreover, the structure of nutrients changed obviously. For Yaling Bay, DIN and DIP increased by about 1.5 and 3.6 times, respectively, but silicate decreased to half after 2003 (Table 2). As nutrients are the material basis for algae growth, the phytoplankton community will show coordinated variation with nutrition structure and concentration. Low silicate could maintain certain diatom diversity but not high diatom biomass. Such nutrition structure variation might be more beneficial to dinoflagellate than diatom in Daya Bay (Wang et al., 2011). Similar trend has been reported in Dapeng Bay in which diatom blooms decreased obviously and dinoflagellate blooms increased according to the decrease of silicate content (Tian et al., 2020). Conversely, in Jiaozhou Bay, a semi-closed bay located in the western area of the Yellow Sea in Shandong Peninsula of China, HABs were almost caused by diatom Skeletonema costatum and Eucampia zoodiacus in accompany with the increase of silicate, DIN, and DIP concentration (Liu et al., 2005; Sun et al., 2011). Consequently, the decrease of silicate in Daya Bay might be an important factor for the increase of dinoflagellate blooms.
Temperature is a vital factor affecting the physiological processes of phytoplankton (Beardall et al., 2009). Many studies have examined that dinoflagellate prefers higher temperature. For example, the genera Ceratium, Protoperidinium and Dinophysis respond quickly to temperature rise, and their growth had high positive correlation with temperature increase in the North Sea (Edwards and Richardson, 2004) and the tropical coastal of south-eastern Arabian Sea (Godhe et al., 2015). In the regions affected by thermal discharge from the power plant, the dominant phytoplankton species shifted from diatoms to dinoflagellates in Xiangshan Port of China (Jiang et al., 2013), Daya Bay (Li et al., 2011), and the coastal waters adjacent to the Changjiang River estuary (Zhou et al., 2017). Especially in areas of higher salinity and higher CO2, increase in temperature leads to an increase in dinoflagellates abundance (Alkawri and Ramaiah, 2010; Keys et al., 2018).
The percentage of dinoflagellate blooms gradually increased to 77% of the total HAB in the DNPP and LNPP operation period in Daya Bay (Fig. 2b). Yaling Bay was the dinoflagellate bloom-prone area with ~30 ℃ sea surface temperature in summer (Huan et al., 2016). Based on the CCA analysis, dinoflagellate abundance was positively correlated with temperature in Yaling Bay (Fig. 3). The trend of correlation between the temperature change and the relative abundance of dinoflagellate was positive (Fig. 3b). In addition, algae growth was also closely related to the water retention time. The seawater retention time in Daya Bay was longer than 3 months, especially in Yaling Bay in summer (Wang et al., 2008, 2009). Furthermore, the thermal effluent will increase the surface seawater temperature to intensify the stratification, which is beneficial to dinoflagellates since dinoflagellates prefer stable waters while diatoms prefer well-mixed waters (Jones and Gowen, 1990; Hinder et al., 2012). Therefore, warm seawater could be the key factor for the increase of dinoflagellate blooms.
4.2 Potential mechanisms for high frequency of S. trochoidea blooms in Daya BayScrippsiella trochoidea was the most important species to induce dinoflagellate blooms in Daya Bay, accounting for about 90% of all dinoflagellate blooms (Fig. 2). S. trochoidea blooms occur in report in water of 25–30 ℃ (Mulholland et al., 2018). As shown in field investigation, the abundance of S. trochoidea increased in 28.2–32.1 ℃. In laboratory incubation, we further found that S. trochoidea grew best around 30 ℃ and quickly reached the exponential growth phase. In addition, Yaling Bay is a shallow and eutrophic inner bay, with a water temperature of about 30 ℃ in summer, which was beneficial to the reproduction and bloom of S. trochoidea (Mulholland et al., 2018).
Furthermore, cysts were important for the appearance of blooms and were also generally considered as the "seed source" of blooms (Wang et al., 2004). In Daya Bay, dinoflagellate cysts were diverse and abundant, and their abundance in Yaling Bay were much higher than that in other areas (Tang, 2018). S. trochoidea bloomed almost every year in Daya Bay since the first observation in 1998. It produced many cysts after bloom and acted as the dominant species across Daya Bay accounting for 51.3%–81.6% of the total cysts, with a high concentration (325 cysts/(g DW)) in Yaling Bay (Wang et al., 2004; Tang, 2018). Generally, the germination of cyst was affected by the length and intensity of dormancy period, temperature, light and dissolved oxygen (Wang et al., 2007). Newly formed cysts required about 25-day dormancy before germination. According to laboratory incubation, weak light intensity could trigger the resting cysts of S. trochoidea germination and the appropriate culture temperature was around 20–25 ℃ (Binder and Anderson, 1986; Qi et al., 1997). To a large extent, illumination (> 0.12 μmol/m2) was enough for its cysts germination by low photon fluency delivered in 1-s exposure (Binder and Anderson, 1986), which could be satisfied considering the transparency in Daya Bay. In summer period, warmer and saltier water from open sea invaded Daya Bay by Guangdong coastal upwelling, and the bottom (about 15 m) temperature is about 20 ℃ (Han and Ma, 1991; Hao et al., 2016). Overall, the environmental conditions of Daya Bay met the needs of cysts of S. trochoidea germination and provided the basis for its bloom.
4.3 General course of S. trochoidea blooms in Daya BayPrior to the onset of blooms, the high tide and strong wind (average daily wind speed > 7 m/s) disturbed the sediment and could resuspend the cysts (Supplementary Table S1). Given that S. trochoidea bloomed almost every year since 1998, cysts were buried shallower than other dinoflagellate cysts and it is easy to resuspend. Under low light intensity and relatively low water temperature (20–25 ℃) (Han, 1998), the cysts of S. trochoidea could germinate rapidly (Qi et al., 1997). The slow water exchange speed gave S. trochoidea enough time for reproduction and gathering at the surface. The sea surface temperature of Yaling Bay was as high as (about 30 ℃) to promote the vegetative cells division. On the other hand, the high temperature seawater and the phytoplankton community affected by thermal discharge stress were taken to the region due to the influence of tide and other hydrodynamic factors. Algae can return to normal physiological state and even have rapid growth in a short time after leaving the transient temperature stimulation (Xie et al., 2015). The stimulating effect of warm drainage from NPPs' was also considered one of the causes of algal blooms (Huang et al., 2002). Coupled with appropriate meteorological conditions (about 9 hours or longer sunlight periods) and weak wind (< 3.3 m/s, light breeze), which provides a stable environment for cells proliferation and gathering together, and then bloom occurs. Furthermore, the degradation of S. trochoidea blooms were accompanied by active encystment (Xiao et al., 2001). After a necessary period of dormancy (about 25 days) (Binder and Anderson, 1986), such cysts could germinate to support another S. trochoidea bloom (Fig. 4).

|
| Fig.4 Conceptual diagram of proposed and hypothesized drivers of S. trochoidea blooms in Daya Bay SST: sea surface temperature; SH: sunlight hours; SSW: south-southwest. |
Previous studies have also shown that the combined effects of warming and eutrophication increased the spatial range and frequency of dinoflagellate blooms (Huang et al., 2002; Zhou et al., 2017), resulting in more frequent and intense dinoflagellate blooms and affecting the function of coastal ecosystems. We analyzed the changes of HABs and the environmental data in Daya Bay in 30 years (1983–2017) combining the meteorological data and laboratory experiments. The proportion of dinoflagellate blooms in Daya Bay increased stepwise, of which over 77% took place after 2011. The bloom-inductive species tended to be singular, as the highest proportion of dinoflagellate blooms were caused by S. trochoidea (89%) in 2003–2010. The higher temperature of seawater might be the key factor for the spatial and temporal HAB patterns in Daya Bay. At the same time, appropriate meteorological and hydrological conditions provided conditions for the occurrence of HABs. Compared with diatom blooms, dinoflagellate blooms are more harmful because they are barely eaten by aquatic organisms and generate toxins (Smayda, 1997). The increase of dinoflagellate blooms in Daya Bay would result in the reduction of fish production and destruction of local ecological environment. Meanwhile, thermal discharge from coast-based NPP would have adverse effects on the coastal ecosystems. Under the theme of global warming, this situation may be worsened. Therefore, it is predictable that dinoflagellates would become more predominant and dinoflagellate blooms would increase in the bay nearby an NPP in coast.
5 DATA AVAILABILITY STATEMENTThe datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
6 ACKNOWLEDGMENTThe reviewers are greatly acknowledged for their constructive comments, which vastly improved this manuscript.
Electronic supplementary materialSupplementary material (Supplementary Table S1) is available in the online version of this article at https://doi.org/10.1007/s00343-020-9082-0.
Adachi R. 1973. Red tide organisms and red tide. Fish Engineering, 9: 31-36.
|
Alkawri A A S, Ramaiah N. 2010. Spatio-temporal variability of dinoflagellate assemblages in different salinity regimes in the west coast of India. Harmful Algae, 9(2): 153-162.
DOI:10.1016/j.hal.2009.08.012 |
Backer L C, McGillicuddy Jr D J. 2006. Harmful algal blooms at the interface between coastal oceanography and human health. Oceanography, 19(2): 94-106.
DOI:10.5670/oceanog.2006.72 |
Beardall J, Stojkovic S, Larsen S. 2009. Living in a high CO2 world: Impacts of global climate change on marine phytoplankton. Plant Ecology & Diversity, 2(2): 191-205.
DOI:10.1080/17550870903271363 |
Binder B J, Anderson D M. 1986. Green light-mediated photomorphogenesis in a dinoflagellate resting cyst. Nature, 322(6080): 659-661.
DOI:10.1038/322659a0 |
Burkholder J M, Glasgow Jr H B. 1997. Pfiesteria piscicida and other Pfiesreria-like dinoflagellates: Behavior, impacts, and environmental controls. Limnology and Oceanography, 42(5): 1 052-1 075.
DOI:10.4319/lo.1997.42.5_part_2.1052 |
Cai Z P, Huang W W, An M, Duan S S. 2009. Coupled effects of irradiance and iron on the growth of a harmful algal bloom-causing microalga Scrippsiella trochoidea. Acta Ecologica Sinica, 29(5): 297-301.
DOI:10.1016/j.chnaes.2009.09.007 |
Edwards M, Richardson A J. 2004. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature, 430(7002): 881-884.
DOI:10.1038/nature02808 |
Gárate-Lizárraga I, Band-Schmidt C J, López-Cortés D J, Socorro Muñetón-Gómez M. 2009. Bloom of Scrippsiella trochoidea (Gonyaulacaceae) in a shrimp pond in the southwestern Gulf of California, Mexico. Marine Pollution Bulletin, 58(1): 145-149.
DOI:10.1016/j.marpolbul.2008.09.016 |
Glibert P M, Berdalet E, Burford M A, Pitcher G C, Zhou M J. 2018. Global Ecology and Oceanography of Harmful Algal Blooms. Springer, Cham. p. 1-461, https://doi.org/10.1007/978-3-319-70069-4.
|
Godhe A, Narayanaswamy C, Klais R, Moorthy K S V, Ramesh R, Rai A, Reddy H R V. 2015. Long-term patterns of net phytoplankton and hydrography in coastal SE Arabian Sea: what can be inferred from genus level data?. Estuarine, Coastal and Shelf Science, 162: 69-75.
DOI:10.1016/j.ecss.2015.03.006 |
Hallegraeff G M. 1992. Harmful algal blooms in the Australian region. Marine Pollution Bulletin, 25(5-8): 186-190.
DOI:10.1016/0025-326X(92)90223-S |
Hameed H A, Saburova M. 2015. First record of Scrippsiella trochoidea (Dinophyceae) in Shatt Al-Arab River (Southern Iraq). Marine Biodiversity Records, 8: e150.
DOI:10.1017/S1755267215001281 |
Han W Y. 1998. Marine Chemistry in South China Sea. Science Press, Beijing. p. 1-289.
(in Chinese)
|
Han W Y, Ma K M. 1991. Study on the process of sea water exchang in Daya Bay. Marine Sciences, (2): 64-67.
(in Chinese with English abstract) |
Hao Y J, Tang D L, Boicenco L, Wang S F. 2016. Environmental ecological response to increasing water temperature in the Daya Bay, Southern China in 1982-2012. Natural Resources, 7(4): 184-192.
DOI:10.4236/nr.2016.74017 |
Hinder S L, Hays G C, Edwards M, Roberts E C, Walne A W, Gravenor M B. 2012. Changes in marine dinoflagellate and diatom abundance under climate change. Nature Climate Change, 2(4): 271-275.
DOI:10.1038/nclimate1388 |
Huan Q L, Pang R S, Zhou Q L, Leng K M. 2016. Variation trends of nitrogen and phosphorus and the relationship with HABs in Shenzhen coastal waters. Marine Environmental Science, 35(6): 908-914.
(in Chinese with English abstract) |
Huang X P, Huang L M, Tan Y H, Zhu L S. 2002. Relationship between red tide and environmental conditions in coastal waters. Marine Environmental Science, 21(4): 63-69.
(in Chinese with English abstract) DOI:10.3969/j.issn.1007-6336.2002.04.015 |
IPCC (Intergovernmental Panel on Climate Change). 2007. Climate Change 2007: the Physical Science Basis. Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge..
|
Jiang Z B, Liao Y B, Liu J J, Shou L, Chen Q Z, Yan X J, Zhu G H, Zeng J N. 2013. Effects of fish farming on phytoplankton community under the thermal stress caused by a power plant in a eutrophic, semi-enclosed bay: induce toxic dinoflagellate (Prorocentrum minimum) blooms in cold seasons. Marine Pollution Bulletin, 76(1-2): 315-324.
DOI:10.1016/j.marpolbul.2013.07.006 |
Jones K J, Gowen R J. 1990. Influence of stratification and irradiance regime on summer phytoplankton composition in coastal and shelf seas of the British Isles. Estuarine, Coastal and Shelf Science, 30(6): 557-567.
DOI:10.1016/0272-7714(90)90092-6 |
Kang D J, Fringer O. 2010. On the calculation of available potential energy in internal wave fields. Journal of Physical Oceanography, 40(11): 2 539-2 545.
DOI:10.1175/2010JPO4497.1 |
Keys M, Tilstone G, Findlay H S, Widdicombe C E, Lawson T. 2018. Effects of elevated CO2 and temperature on phytoplankton community biomass, species composition and photosynthesis during an experimentally induced autumn bloom in the western English Channel. Biogeosciences, 15(10): 3 203-3 222.
DOI:10.5194/bg-15-3203-2018 |
Kim Y O, Han M S. 2000. Seasonal relationships between cyst germination and vegetative population of Scrippsiella trochoidea (Dinophyceae). Marine Ecology Progress Series, 204: 111-118.
DOI:10.3354/meps204111 |
Knap A H, Michaels A, Close A R, Ducklow H, Dickson A G. 1996. Protocols for the Joint Global Ocean Flux Study (JGOFS) Core Measurements. UNESCO, Bergen.
|
Kumar P S, Kumaraswami M, Rao G D, Ezhilarasan P, Sivasankar R, Rao V R, Ramu K. 2018. Influence of nutrient fluxes on phytoplankton community and harmful algal blooms along the coastal waters of southeastern Arabian Sea. Continental Shelf Research, 161: 20-28.
DOI:10.1016/j.csr.2018.04.012 |
Li L, Lü S H, Cen J Y. 2019. Spatio-temporal variations of Harmful algal blooms along the coast of Guangdong, Southern China during 1980-2016. Journal of Oceanology and Limnology, 37(2): 535-551.
DOI:10.1007/s00343-019-8088-y |
Li T, Liu S, Huang L M, Huang H, Lian J S, Yan Y, Lin S J. 2011. Diatom to dinoflagellate shift in the summer phytoplankton community in a bay impacted by nuclear power plant thermal effluent. Marine Ecology Progress Series, 424: 75-85.
DOI:10.3354/meps08974 |
Licea S, Zamudio M E, Luna R, Okolodkov Y, Gómez-Aguirre S. 2004. Toxic and harmful dinoflagellates in the southern Gulf of México. In Steidinger K A, Landsberg J H, Tomas C R, Vargo G A eds. Harmful Algae 2002. Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography and Intergovernmental Oceanographic Commission of UNESCO, St. Petersburg. p. 380-382.
|
Lin G H, Dong Y R, Li K Q, Wang S P, Wang X L. 2017. The method to identity the key control factor of red tide: a case study of Bohai Sea. Periodical of Ocean University of China, 47(12): 88-96.
(in Chinese with English abstract) DOI:10.16441/j.cnki.hdxb.20160287 |
Lin J N, Yan T, Zhang Q C, Wang Y F, Liu Q, Zhou M J. 2014. In situ detrimental impacts of Prorocentrum donghaiense blooms on zooplankton in the East China Sea. Marine Pollution Bulletin, 88(1-2): 302-310.
DOI:10.1016/j.marpolbul.2014.08.026 |
Lin X, Zhang H, Huang B Q, Lin S J. 2012. Alkaline phosphatase gene sequence characteristics and transcriptional regulation by phosphate limitation in Karenia brevis (Dinophyceae). Harmful Algae, 17: 14-24.
DOI:10.1016/j.hal.2012.02.005 |
Lin Z X, Liang S H. 2002. Study and forecasting on red tide caused by marine physical factors. Mar. Environ. Sci., 21(2): 1-5.
(in Chinese with English abstract) |
Liu D Y, Sun J, Zou J Z, Zhang J. 2005. Phytoplankton succession during a red tide of Skeletonema costatum in Jiaozhou Bay of China. Marine Pollution Bulletin, 50(1): 91-94.
DOI:10.1016/j.marpolbul.2004.11.016 |
Lu D D, Goebel J. 2001. Five red tide species in genus Prorocentrum including the description of Prorocentrum donghaiense Lu SP. nov. from the east china sea. Chinese Journal of Oceanology and Limnology, 19(4): 337-344.
DOI:10.1007/BF02850738 |
Mulholland M R, Morse R, Egerton T, Bernhardt P W, Filippino K C. 2018. Blooms of dinoflagellate mixotrophs in a lower Chesapeake Bay tributary: carbon and nitrogen uptake over diurnal, seasonal, and interannual timescales. Estuaries and Coasts, 41(6): 1 744-1 765.
DOI:10.1007/s12237-018-0388-5 |
Qi Y Z, Zheng L, Wang R. 1997. The life cycle of Scrippsiella trochoidea and its physiol-ecological control. Oceanologia et Limnologia Sinica, 28(6): 588-593.
(in Chinese with English abstract) |
Qian H L, Liang S, Qi Y Z. 2000. Study of the characteristics and the causes of formation on the red tides in coastal Guangdong sea. Ecologic Science, 19(3): 8-16.
(in Chinese with English abstract) DOI:10.3969/j.issn.1008-8873.2000.03.002 |
Shin H H, Li Z, Yoon Y H, Oh S J, Lim W A. 2017. Formation and germination of temporary cysts of Cochlodinium polykrikoides Margalef (Dinophyceae) and their ecological role in dense blooms. Harmful Algae, 66: 57-64.
DOI:10.1016/j.hal.2017.05.002 |
Smayda T J. 1997. Harmful algal blooms: their ecophysiology and general relevance to phytoplankton blooms in the sea. Limnology and Oceanography, 42(5): 1 137-1 153.
DOI:10.4319/lo.1997.42.5_part_2.1137 |
Smayda T J, Reynolds C S. 2003. Strategies of marine dinoflagellate survival and some rules of assembly. Journal of Sea Research, 49(2): 95-106.
DOI:10.1016/S1385-1101(02)00219-8 |
Sun J, Ning X R. 2005. Marine phytoplankton specific growth rate. Advances in Earth Science, 20(9): 939-945.
(in Chinese with English abstract) DOI:10.3321/j.issn:1001-8166.2005.09.003 |
Sun X X, Sun S, Zhao Z X, Shen Z L. 2011. Long-term changes in nutrient concentration and structure in the Jiaozhou Bay. Oceanologia et Limnologia Sinica, 42(5): 662-669.
(in Chinese with English abstract) |
Tang D L, Kester D R, Wang Z D, Lian J S, Kawamura H. 2003. AVHRR satellite remote sensing and shipboard measurements of the thermal plume from the Daya Bay, nuclear power station, China. Remote Sensing of Environment, 84(4): 506-515.
DOI:10.1016/S0034-4257(02)00149-9 |
Tang Y N. 2018. Distribution of Dinoflagellate Cysts in the Daya Bay and its Relations to Environmental Factors. University of Chinese Academy of Sciences, p. 1-67. (in Chinese with English abstract)
|
Tang Y Z, Gobler C J. 2015. Sexual resting cyst production by the dinoflagellate Akashiwo sanguinea: a potential mechanism contributing to the ubiquitous distribution of a harmful alga. Journal of Phycology, 51(2): 298-309.
DOI:10.1111/jpy.12274 |
Tian Y, Li T, Hu S M, Xie X D, Liu S. 2020. Temporal and spatial characteristics of Harmful algal blooms in Guangdong coastal area. Marine Environmental Science, 39(1): 1-8.
(in Chinese with English abstract) |
Townhill B L, Tinker J, Jones M, Pitois S, Creach V, Simpson S D, Dye S, Bear E, Pinnegar J K. 2018. Harmful algal blooms and climate change: exploring future distribution changes. ICES Journal of Marine Science, 75(6): 1 882-1 893.
DOI:10.1093/icesjms/fsy113 |
Wang C, Lin J, Chen P M, Zhang S Y. 2008. Numerical simulation on water exchange in Daya Bay. South China Fisheries Science, 4(4): 8-15.
(in Chinese with English abstract) DOI:10.3969/j.issn.2095-0780.2008.04.002 |
Wang C, Lin J, Chen P M, Zhang S Y. 2009. Numerical simulation of annual average wind's impact on water exchange in Daya Bay. Journal of Shanghai University, 18(3): 351-358.
(in Chinese with English abstract) |
Wang Y S, Sun C C, Lou Z P, Wang H L, Mitchell B G, Wu M L, Sun Z X. 2011. Identification of water quality and benthos characteristics in Daya Bay, China, from 2001 to 2004. Oceanological and Hydrobiological Studies, 40(1): 82-95.
DOI:10.2478/s13545-011-0009-4 |
Wang Z H, Qi Y Z, Jiang T J, Xu Z N. 2004. Vertical distribution of dinoflagellate resting cysts in recent sediments from Daya Bay, the south china sea. Acta Hydrobiologica Sinica, 28(5): 504-510.
(in Chinese with English abstract) DOI:10.3321/j.issn:1000-3207.2004.05.008 |
Wang Z H, Qi Y Z, Yang Y F. 2007. Cyst formation: an important mechanism for the termination of Scrippsiella trochoidea (Dinophyceae) bloom. Journal of Plankton Research, 29(2): 209-218.
DOI:10.1093/plankt/fbm008 |
Wells M L, Trainer V L, Smayda T J, Karlson B S O, Trick C G, Kudela R M, Ishikawa A, Bernard S, Wulff A, Anderson D M. 2015. Harmful algal blooms and climate change: learning from the past and present to forecast the future. Harmful Algae, 49: 68-93.
DOI:10.1016/j.hal.2015.07.009 |
Xiao W P, Liu X, Irwin A J, Laws E A, Wang L, Chen B Z, Zeng Y, Huang B Q. 2018. Warming and eutrophication combine to restructure diatoms and dinoflagellates. Water Research, 128: 206-216.
DOI:10.1016/j.watres.2017.10.051 |
Xiao Y Z, Qi Y Z, Wang Z H, Lü S H. 2001. The relationship between Scrippsiella trochoidea red tide and cysts in the Daya Bay. Marine Sciences/Monthly, 25(9): 50-54.
(in Chinese with English abstract) DOI:10.3969/j.issn.1000-3096.2001.09.017 |
Xiao Y Z, Wang Z H, Chen J F, Lü S H, Qi Y Z. 2003. Seasonal dynamics of dinoflagellate cysts in sediments from Daya Bay, the South China Sea its relation to the bloom of Scrippsiella trochoidea. Acta Hydrobiologica Sinica, 27(4): 372-377.
(in Chinese with English abstract) DOI:10.3321/j.issn:1000-3207.2003.04.008 |
Xie Y H, Li T, Jian W J, Hu S M, Tian Y, Liu S. 2015. Influence of ocean warming on the community structure and photosynthetic efficiency of phytoplankton in Daya Bay. Journal of Tropical Oceanography, 34(2): 24-31.
(in Chinese with English abstract) DOI:10.3969/j.issn.1009-5470.2015.02.004 |
Xu N, Lü S H, Chen J F, He L S, Xie L C, Qi Y Z. 2004. The influence of water temperature and salinity on the growth of Scrippsiella trochoidea. Marine Environmental Science, 23(3): 36-38.
(in Chinese with English abstract) DOI:10.3969/j.issn.1007-6336.2004.03.011 |
Xu N, Pang S J. 2012. Molecular identification and culture trials of three marine microalgae. Marine Sciences, 36(10): 13-25.
(in Chinese with English abstract) |
Yin K D, Song X X, Liu S, Kan J J, Qian P Y. 2008. Is inorganic nutrient enrichment a driving force for the formation of red tides?: A case study of the dinoflagellate Scrippsiella trochoidea in an embayment. Harmful Algae, 8(1): 54-59.
DOI:10.1016/j.hal.2008.08.004 |
Yu J, Tang D L, Oh I S, Yao L J. 2007. Response of harmful algal blooms to environmental changes in Daya Bay, China. Terrestrial, Atmospheric and Oceanic Sciences Journal, 18(5): 1 011-1 027.
DOI:10.3319/TAO.2007.18.5.1011(Oc) |
Zhou Z X, Yu R C, Zhou M J. 2017. Resolving the complex relationship between harmful algal blooms and environmental factors in the coastal waters adjacent to the Changjiang River estuary. Harmful Algae, 62: 60-72.
DOI:10.1016/j.hal.2016.12.006 |