 2023, Vol. 41
2023, Vol. 41Institute of Oceanology, Chinese Academy of Sciences
Article Information
- CHEN Yuzhen, ZHU Huilan, GUO Yuqing
- A new species and a new record of free-living marine nematode of genus Diplolaimelloides (Monhysteridae) from Futian mangrove reservation in Shenzhen, China
- Journal of Oceanology and Limnology, 41(1): 314-326
- http://dx.doi.org/10.1007/s00343-021-1206-7
Article History
- Received Jul. 1, 2021
- accepted in principle Sep. 13, 2021
- accepted for publication Dec. 13, 2021




The family Monhysteridae is one of the most successful families within the free-living nematodes, it displays a large number of individuals and species in marine, brackish, limnic, and terrestrial biotopes (Lorenzen, 1978; Fonseca and Decraemer, 2008). Species of Diplolaimelloides Meyl, 1954 can be found in salty (Timm, 1966) and brackish water (Tsalolikhin, 1985). Some species can be cultured easily in laboratories, for example D. meyli Timm, 1961 and D. brucei Hopper, 1970, showing activity within a wide salinity range (Allgén, 1933; Warwick, 1981; Moens and Vincx, 2000; de Ward and Russo, 2009).
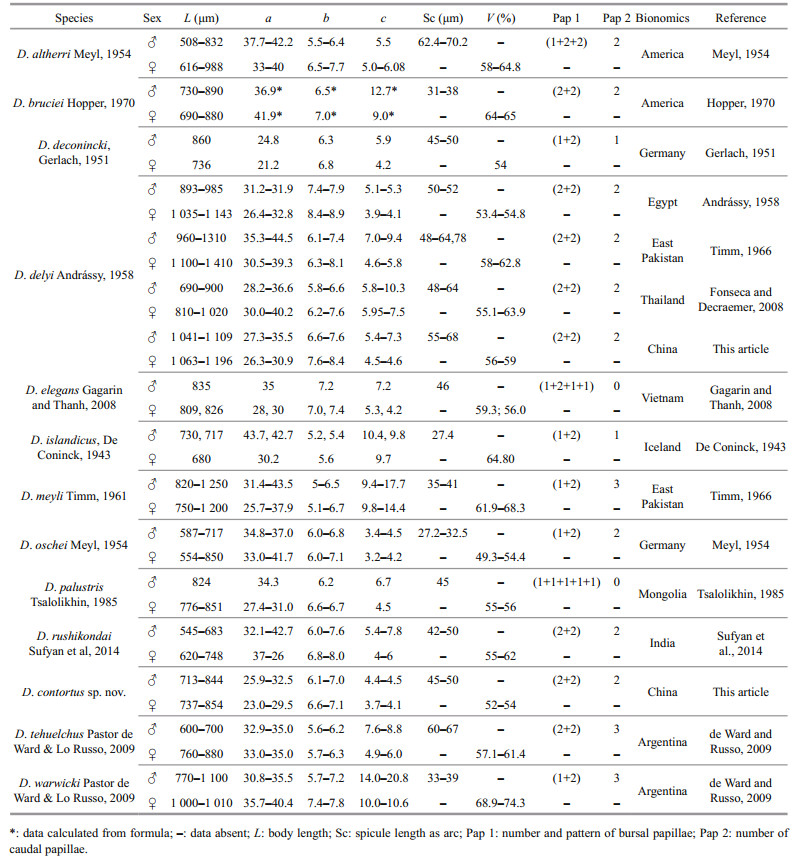
The genus Diplolaimelloides was erected by Meyl (1954) for Diplolaimella species in which the male has a genital bursa with fine genital papillae. De Coninck (1965) set up a new subfamily, the Diplolaimelloidinae, to embrace the species of Diplolaimelloides, based on the possession of a genital bursa by the males. Timm (1966) considered that it seems best to keep Diplolaimelloides and Diplolaimella in a single subfamily, since the morphological relationships between these two genera is quite apparent, and they were found in the same or similar habitats. The taxonomy of Diplolaimelloides has been comprehensively revised many times (Timm, 1966; Gerlach and Riemann, 1973; Fonseca and Decraemer, 2008; de Ward and Russo, 2009). Characters such as the presence or absence of ocelli or amphids, distance of ocelli or amphidial fovea from anterior end, tail length and de Man's ratios have been considered to be the most important feature to separate the species of Diplolaimelloides (Fonseca and Decraemer, 2008; de Ward and Russo, 2009), and the number and arrangement pattern of bursal and caudal papillae, morphology of spicules and gubernaculum can help in better diagnosis of species (Sufyan et al., 2014).
One new species and one new record of the genus of Diplolaimelloides were found during a preliminary survey of the meiofauna in mangrove mudflats of Shenzhen, China, and are described in the present paper.
2 MATERIAL AND METHODSediment samples were collected from Futian mangrove mudflats in Shenzhen, China (22.52°N, 114.01°E), and fixed with 5% formalin in filtered seawater immediately. Nematodes were extracted from the sediment by decantation and/or Ludox centrifugation in the laboratory, and stained with rose Bengal for more than 24 h in the laboratory (Warwick et al., 1998). Then nematodes were transferred to a cavity block full with a solution containing, by volume, 5% glycerol, 5% pure ethanol, and 90% freshwater, to let ethanol slowly evaporate, and then mounted nematode in glycerol on permanent slides (McIntyre and Warwick, 1984). The observations were made from glycerin mounts using differential contrast microscopy (NIKON 80i). Drawings were performed using a camera lucida.
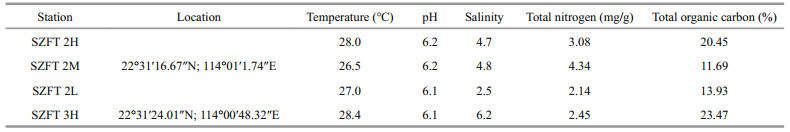
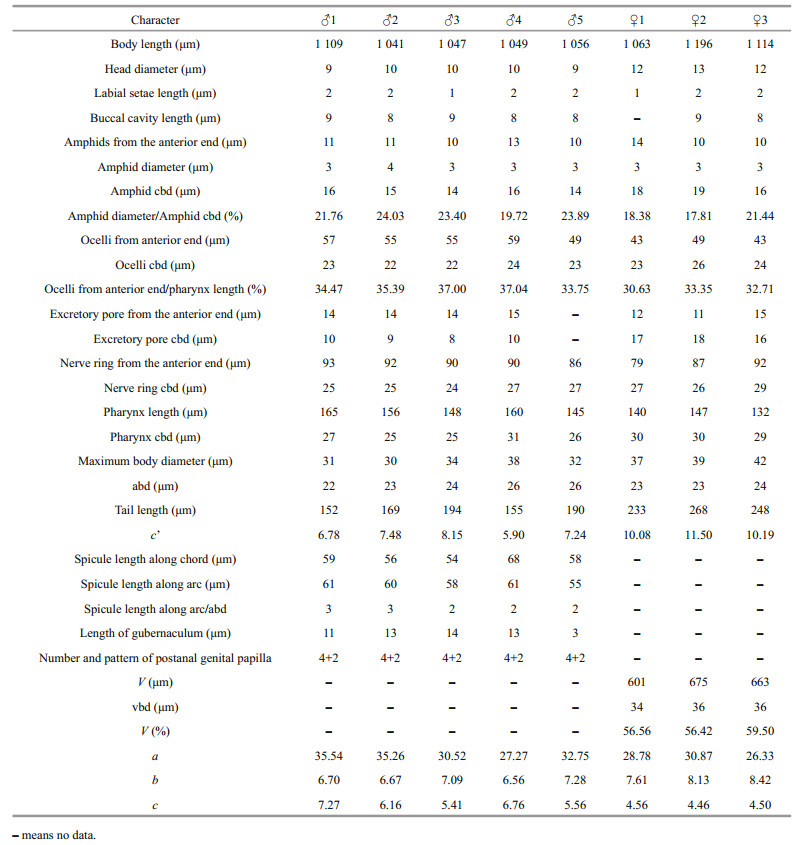
Measurements are in μm. Abbreviations are as follows: a: body length/maximum body diameter; b: body length/pharynx length; c: body length/tail length; abd: anal body diameter; cbd: corresponding body diameter; vbd: vulval body diameter; c': tail length/abd; V: vulva from the anterior end; V(%): position of vulva as % of body length from anterior end, that is, V/body length. All holotypes and paratypes are deposited in the Institute of Oceanology, Chinese Academy of Sciences, Qingdao, China.
3 RESULT AND DISCUSSION 3.1 SystematicFamily Monhysteridae de Man, 1876
Genus Diplolaimelloides Meyl, 1954
Diagnosis (Modified from Fonseca and Decraemer, 2008) Outer labial and cephalic sensillae small, setiform. Buccal cavity with two chambers. Amphidial fovea cryptospiral, i.e. ventrally wound spiral with a broken or circular aspect, placed 1–2 times of head diameter from anterior end. Ventral gland mostly present, opening anterior to nerve ring (mid-pharynx). Ocelli usually present, varying in position from anterior end. Vulva located in the mid-body region of long-tailed species, and more posterior in shorter-tailed ones. Spicules slender, arcuate to almost straight, varying in length from 1.8 to 4.4 abd. Gubernaculum poorly sclerotized, without dorsal apophysis. Male with genital bursa, which encompassing the anterior portion of the tail and supported by several pairs of fine postcloacal papillae. Tail with posterior section cylindrical. Reproductive mode: bisexual (Fonseca and Decraemer, 2008).
There are 12 valid species in this genus up to now (Gerlach and Riemann, 1973; de Ward and Russo, 2009; Sufyan et al., 2014; Electronic database: http://nemys.ugent.be; http://www.marinespecies.org/index.php), in which D. deconincki (Gerlach, 1951) and D. islandicus (De Coninck, 1943) were first described as species of Diplolaimella, and then transferred to Diplolaimelloides. Two species Diplolaimelloides longispicula and Diplolaimella monhysteroides are considered to be inquirendum. Jacobs (1987) gave a checklist for the genus recognized valid species, including a new species D. longispicula Jacobs, 1987, for D. monhysteroides Allgén, 1929 sensu Allgén, 1930. Since D. longispicula Jacobs, 1987 was described based on a single male which not bursa or papillae were described or illustrated, it is considered to be inquirendum (Fonseca and Decraemer, 2008). Diplolaimella monhysteroides was described by Allgén, 1929 sensu Allgén, 1930 as the type species of Diplolaimella. Timm (1966) considerd that even the female of D. monhysteroides belonged to Diplolaimella rather than Diplolaimelloides as it is presently understood, the body of Diplolaimella is generally thinner than that of Diplolaimelloides and in this respect D. monhysteroides clearly conforms to Diplolaimella. However, the more posterior position of the vulva (64%) is characteristic of Diplolaimelloides, and the spicules and shape of the tail are characteristic of Diplolaimelloides but the thin bursa and thickening of the wall of the rectum were overlooked. Therefore, D. monhysteroides must be regarded as a species dubia.
With the exception of D. longispicula and D. monhysteroides, all valid species are listed below.
List of valid Diplolaimelloides species
D. altherri Meyl, 1954
D. bruciei Hopper, 1970
D. deconincki (Gerlach, 1951) syn, Diplolaimella deconincki Gerlach, 1951
D. delyi Andrássy, 1958
D. elegans Gagarin & Thanh, 2008
D. islandicus (De Coninck, 1943) syn.
Diplolaimella islandicus (De Coninck, 1943)
D. meyli Timm, 1961
D. oschei Meyl, 1954
D. palustris Tsalolikhin, 1985
D. rushikondai Sufyan et al, 2014
D. tehuelchus Pastor de Ward, 2009
D. warwicki Pastor de Ward, 2009
List of inquirendum Diplolaimelloides species
D. longispicula Jacobs, 1987 syn. Diplolaimella monhysteroides apud Allgén, 1930.
D. monhysteroides Allgén, 1930.
3.2 Description of new speciesDiplolaimelloides contortus sp. nov. (Figs. 1–2)

|
| Fig.1 Diplolaimelloides contortus sp. nov. a. lateral view of male body; b. lateral view of female tail region; c. lateral view of male tail region; d. lateral view of male head end; e. lateral view of female head end; f. lateral view of female anterior part; g. lateral view of copulatory structure. Scale bars: a=125 μm; b–f=50 μm; g=10 μm. |

|
| Fig.2 Diplolaimelloides contortus sp. nov. a. lateral view of male head end, showing amphid; b. lateral view of male body part, showing spicule; c. lateral view of male body part, showing spicule and bursal papillae; d. lateral view of male total body; e. lateral view of male tail region; f. lateral view of male body part, showing bursa; g. lateral view of female head end, showing amphid, ocelli, and pharynx; h. lateral view of female body part, showing ovary; i. lateral view of female head end, showing amphid; j. lateral view of female head end, showing buccal cavity; k. lateral view of female total body; l. lateral view of female tail region. Scale bar showing in (l): a–c, f, g, i, j=20 μm; e, h=50 μm; k, l=100 μm; d=200 μm. |
Four males and three females were collected from Station SZFT in July, 2020.
Holotype: ♂1 on slide number SZFT202007 3H114. Paratypes: ♂2 on slide number SZFT202011 2H120, ♂3 on slide number SZFT202007 2H120, ♂4 on slide number SZFT202007 3H115, ♀1 on slide number SZFT202007 3H112, ♀2 and ♀3 on slide number SZFT202007 3H107.
3.2.2 Type locality and habitatAll specimens were collected from a mangrove forest in the mangrove reservation of Shenzhen, China. Characteristics of surface sediments of sampling stations are shown in Table 1.
The specific name is from Latin (contortus = whirling) and refers to the intricate organization of the copulatory structure and accessory organ.
3.2.4 MeasurementMorphological characteristics were observed and measured under a differential contrast microscopy (NIKON 80i) (Table 2).

Body slender and cylindrical, maximum body diameter 25–33 μm. Cuticle smooth. Inner labial sensilla organs not detected, six outer labial and four cephalic sensilla setiform, about 1–2 μm. Head diameter 7–9 μm, with clear area at anterior. Buccal cavity lightly sclerotized at anterior and posterior chambers (varying degrees of sclerotization in different specimens), but the junction of the two chambers sclerotized more heavily in each specimen; no denticles in buccal cavity. Amphids single spiral, 2.9–3.6 μm in diameter or 23%–31% of corresponding body diameter, located 9–10 μm from anterior end, or 1.02–1.28 times of head diameters. Ocelli 35–49 μm from anterior end, or 32%–41% of pharynx length from anterior end. Pharynx cylindrical, 0.14–0.16 times of body length, gradually enlarging in posterior portion but without bulb. Nerve ring located at 44%–59% of pharynx length from anterior end. Cardia distinct between pharynx and intestine.
Male monorchic, testis outstreched on the right side of intestine. Tail slender, 10.0–11.4 abd, with proximal about 25% of length conical and gradually going over into distal cylindrical portions. Bursa 50–60-μm long along arc, with four pairs of post-cloacal papillae arranged in (2+2) pattern, two midway on bursa, along bursal arc about 11–13 μm and 14–16 μm posterior to cloaca, respectively, and two near posterior extremities, along bursal arc about 27–31 μm and 30–33 μm posterior to cloaca, respectively. Two additional pairs of papillae also found on the tail, one pair at junction of conical and filiform parts of tail and one pair of papillae near anterior of filiform part of tail, enclosed in a thin projection of bursa. Spicules complicated in shape, twisted and curved at middle portion, the diagonal list that can be observed in all species of this group is here developed into a lateral plate connecting proximal and distal end of spicule. Gubernaculum wedge-shaped, with distal dorsal ridge well cuticularized, proximal dorsal ridge obscure sleeve-like, proximal end wrap the spicule.
Female reproductive system mono-prodelphic. Ovary outstretched, present on right side of intestine. Vulva located next to mid-body region (52%–54% of body length). Tail long filiform, 13.1–13.6 abd.
3.2.6 DiagnosisDiplolaimelloides contortus sp. nov. is characterized by the combination of the following characters: ocelli present; four pairs of post-cloacal papillae arranged in (2+2) pattern, two midway on bursa and two near posterior extremities; Spicules and gubernaculum complicated in shape; vulva located at 52%–54% of body length; male tail slender, 10.0–11.4 abd, and female tail long filiform, 13.1–13.6 abd. From all valid species reviewed, this new species resembles D. delyi Andrássy, 1958, D. bruciei Hopper, 1970, D. rushikondai Sufyan et al., 2014, and D. tehuelchus Pastor de Ward & Lo Russo, 2009 by having four papillae on bursa 2+2 arrangement. However, D. contortus can be easily distinguished from them by the shape and length of spicule, the more anterior position of vulva, the relatively longer tail and the smaller value of a, b, and c.
3.3 Description of new recordDiplolaimelloides delyi Andrássy, 1958 (Figs. 3–4; Table 3)

|
| Fig.3 Diplolaimelloides delyi Andrássy, 1958 a. lateral view of male tail region; b. lateral view of male head end; c. lateral view of female anterior part; d. lateral view of female tail region; e. lateral view of female head end; f. lateral view of male body. Scale bars: 50 μm. |

|
| Fig.4 Diplolaimelloides delyi Andrássy, 1958 a. lateral view of male head end, showing buccal cavity; b. lateral view of male head end, showing amphid and ocelli; c. lateral view of female head end, showing ocelli; d. lateral view of female tail region; e. lateral view of male tail region; f. lateral view of female body part, showing vulva; g. lateral view of male body part, showing spicule and gubernaculum; h. lateral view of male total body; i. lateral view of female total body; j. lateral view of male body part, showing bursa. Scale bar showing in (j): a, b, c, f, g, j=20 μm; e=50 μm; d, h=100 μm; i=200 μm. |
Five males and three females were collected from Station SZFT in July 2020.
♂1 on slide number SZFT202007 3H102. ♂2 and ♀1 on slide number SZFT202007 3H105, and ♂3 on slide number SZFT202007 2M104, ♂4 on slide number SZFT202007 2L107, ♂5 on slide number SZFT202007 2M103, ♀2 on slide number SZFT202007 3H120, ♀3 on slide number SZFT202007 2M115.
3.3.2 Type locality and habitatAll specimens were collected from a mangrove forest in the mangrove reservation of Shenzhen, China. Characteristics of surface sediments of sampling stations are shown in Table 1.
3.3.3 MeasurementMorphological characteristics were observed and measured under a differential contrast microscopy (NIKON 80i) (Table 3).
3.3.4 DescriptionBody slender and cylindrical, maximum body diameter 30–42 μm. Cuticle smooth. Inner labial sensilla organs not detected, six outer labial and four cephalic sensilla setiform, about 1–2 μm. Head diameter 9–10 μm in male and 12–13 μm in female, with clear area at anterior. Buccal cavity lightly sclerotized at anterior and posterior chambers (varying degrees of sclerotization in different specimens), but the junction of the two chambers sclerotized more heavily in each specimen; no dorsal tooth in posterior chamber. Amphids single spiral, 3–4 μm in diameter or 18%–24% of corresponding body diameter, located 10–14 μm from anterior end, or 0.8–1.3 head diameters. Ocelli consisting of small lens and large red pigment mass, located 49–59 μm from anterior end in male, 43–49 μm in female, or 34%–37% of pharynx length from anterior end in male and 31%–33% in female. Excretory pore located at amphids level or a little posterior to amphids. Pharynx 0.14–0.15 times of body length in male and 0.12–0.13 times of body length in female, cylindrical, gradually enlarging in posterior portion but without bulb. Nerve ring located at 56%–69% of pharynx length from anterior end. Cardia distinct between pharynx and intestine.
Male monorchic, testis outstreched on the right side of intestine. Tail divisible into two parts, anterior conical part about 0.25 times of tail length, posterior part filiform. Bursa 61–85-μm long along arc, with four pairs of post-cloacal papillae arranged in (2+2) pattern, two midway on bursa, along bursal arc about 6–9 μm and 8–10 μm posterior to cloaca, respectively, and two near posterior extremities, along bursal arc about 24–30 μm and 25–32 μm posterior to cloaca, respectively. Two additional pairs of papillae also found on the tail, one dorso-lateral pair just below bursa and one lateral pair about two anal body diameters posterior to cloaca. Spicules 54–68-μm along arc, spicule tip heavily sclerotized on ventral side but about half of ventral side of blade unsclerotized and twisted outward. Gubernaculum small, wedge-shaped, proximal end raised to form a groove.
Female reproductive system mono-prodelphic. Ovary outstretched, present on right side of intestine. Vulva at 56.4%–59.5% of body length. Tail long filiform, 10.1–11.5 times of anal body diameter.
3.3.5 DiagnosisDiplolaimelloides delyi was described by Andrássy in 1958 and several redescriptions have been given (Andrássy, 1958; Timm, 1966). According to the description of Timm (1966), it is characterized by tail long; spicules distinctly cephalate and bearing light transverse striations, spicule tip heavily sclerotized on ventral side but most of ventral side of blade unsclerotized and twisted outward; gubernaculum consisting of a simple thickening of posterior rectal lining; genital bursa with four pairs of postanal genital papillae arranged in (2+2), two pairs of caudal papillae, one pair at junction of conical and filiform parts of tail and one ventro-lateral pair of papillae near anterior of filiform part of tail. Our specimens are much conformed to these characteristics.
Compared with the original description by Andrássy in 1958, our specimens differs from it in larger body size of male (1 041–1 109 μm vs. 893–985 μm), shorter tail (c=5.4–7.3 in male and 4.5–4.6 in female vs. 5.1–5.3 in male and 3.9–4.1 in female), posterior situation of vulva (V=56.4%–59.5% vs. 53.4%–54.8%) (Andrássy, 1958). But our specimens very much conform to the redescription to specimens from East Pakistan by Timm in 1966, except the tail shorter in female (c=4.5–4.6 in female vs. c=4.6–5.8), the pre-anal seta absent (absent vs. one tiny mid-ventral pre-anal seta), the spicules did not cephalate and without transverse striations (Timm, 1966).
3.4 DiscussionThere are 12 valid species in this genus have been described up to now (Gerlach and Riemann, 1973; de Ward and Russo, 2009; Sufyan et al., 2014; Electronic database: http://nemys.ugent.be; http://www.marinespecies.org/index.php), Table 4 shows the differentiating characters of all known Diplolaimelloides species. Dichotomous key in this genus have been revised several times. We revise the key base on de Ward and Russo (2009) which is considering male characters only, including all known valid species as well as the new described species. Figure 5 shows the pictorial key to 12 valid species as well as the newly described species of the genus Diplolaimelloides.

|
| Fig.5 Pictorial key for genus Diplolaimelloides |
Key to species of the genus Diplolaimelloides
1. Bursal papillae in pattern (2+2)............... 2
- Bursal papillae in other patterns............... 6
2. Male tail conical at anterior, filamentous in posterior part, filliform part more than half of tail length..............3
- Male tail elongate-conoid, 2.0-3.5 abd, posterior two-fifths cylindroid..............D. bruciei Hopper, 1970
3. Male tail length less than 5 abd........................D. tehuelchus de Ward & Russo, 2009
- Male tail length more than 5 abd.............. 4
4. Ocelli absent.......................................................D. rushikondai Sufyan et al, 2014
- Ocelli present........................................... 5
5. Spicules distinctly cephalate and bearing light transverse striations; spicule tip heavily sclerotized on ventral side but most of ventral side of blade unsclerotized and twisted outward..........................D. delyi Andrássy, 1958 (Timm, 1966)
- Spicules twisted and curved at middle portion, from where developed into a lateral plate connecting proximal and distal end of spicule............................D. contortus sp. nov.
6. Male without caudal papillae.................. 7
- Male with caudal papillae....................... 8
7. Bursal papillae in pattern (1+1+1+1+1)..........D. palustris Tsaliolikhin, 1985
- Bursal papillae in pattern (1+2+1+1)...............D. elegans Gagarin & Thanh, 2008
8. With 3 papillae in bursa............................ 9
- With 5 papillae in bursa....................................D. altherri Meyl, 1954
9. Male tail relatively long, length 14 abd................D. oschei Meyl, 1954
- Male tail relatively short, length less than 10 abd................................................................................ 10
10. With one male tail papilla...................... 11
- With more male tail papillae.................... 12
11. Spicules shorter than 30 μm..........................D. islandicus (De Coninck, 1943)
- Spicules longer than 30 μm...............................D. deconincki (Gerlach 1951)
12. Spicules angular and long gubernaculum …....D. warwicki de Ward & Russo, 2009
- Spicules arcuate and short gubernaculum.............D. meyli Timm, 1961
4 DATA AVAILABILITY STATEMENTThe data generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
5 ACKNOWLEDGMENTThe authors thank Futian-CityU Mangrove Research & Development Centre for the help in sampling.
Allgén C A. 1933. Zur sammel-, konservierungs- und präparationstechnik freilebender mariner nematoden. Kongelige Norske Vidensh Selskabs Forhandlinger, 6: 97-100.
|
Andrássy I. 1958. Nematoden aus ägyptischen Gewässern (Ergebnisse der zoologischen Aufsammlungen des Ungarischen Naturwissensch. Museums in Ägypten im Jahre 1957). Annales Historico-Naturales Musei Nationalis Hungarici, 9(50): 135-150.
|
De Coninck L A. 1943. Sur Quelques espèces nouvelles de Nématodes libres des eaux et des terres saumâtres de l'Islande. Biologisch Jaarboek, 10: 193-220.
|
De Coninck L A. 1965. Classe des nématodes—systématique des nématodes et sous-classe des adenophorea. In: Grassé P P ed. Traité de Zoologie. Masson, Paris. p. 1-731.
|
de Man J G. 1876. Onderzoekingen over vrij in de aarde levende Nematoden. Tijdschrift der Nederlandsche dierkundige Vereeniging, 2: 78-196.
|
de Ward C T P, Russo V L. 2009. Distribution of Diplolaimella and Diplolaimelloides species from Patagonian lagoons and coastal waters (Nematoda: Monhysteridae), Chubut and Santa Cruz provinces (Argentina). Journal of the Marine Biological Association of the United Kingdom, 89(4): 711-718.
DOI:10.1017/S0025315409000198 |
Fonseca G, Decraemer W. 2008. State of the art of the free-living marine Monhysteridae (Nematoda). Journal of the Marine Biological Association of the United Kingdom, 88(7): 1371-1390.
DOI:10.1017/S0025315408001719 |
Gagarin V G, Thanh N V. 2008. Four new species of monhysterids (Nematoda: Monhysterida) from mangroves of the Mekong river estuaries of Vietnam. Academia Journal of Biology, 30(4): 16-25.
DOI:10.15625/0866-7160/v30n4.5446 |
Gerlach S A, Riemann F. 1973. The bremerhaven checklist of aquatic nematodes. A catalogue of nematoda adenophorea excluding the dorylaimida. Veröffentlichungen des Instituts für Meeresforschung in Bremerhaven, 4(S1): 1-404.
|
Gerlach S A. 1951. Freilebende nematoden aus der verwandtschaft der gattung Theristus. Zoologische Jahrbücher, 80: 379-406.
|
Hopper B E. 1970. Diplolaimelloides bruciei n. sp. (Monhysteridae: Nematoda), prevalent in marsh grass, Spartina alterniflora Loisel. Canadian Journal of Zoology, 48(3): 573-575.
DOI:10.1139/z70-095 |
Jacobs L J. 1987. A Checklist of the Monhysteridae (Nematoda, Monhysterida). Randse Afrikaanse Universiteit, Johannesburg, South Africa.
|
Lorenzen S. 1978. The system of the Monhysteroidea (Nematodes): a new approach. Zoologischer Jahrbucher zur Systematik, 105(4): 515-536.
|
McIntyre A D, Warwick R M. 1984. Meiofauna techniques. In: Holme N A, McIntyre A D eds. Methods for the Study of Marine Benthos. Blackwell Scientific Publications, Oxford, Britain, p. 217-244.
|
Meyl A H. 1954. Die Fadenwürmer (Nematoda) einiger Salzstellen südöstlich von Braunschweig. Abhandlungen der Braunschweigischen wissenschaftlichen Gesellschaft, 6: 84-106.
|
Moens T, Vincx M. 2000. Temperature and salinity constraints on the life cycle of two brackish-water nematode species. Journal of Experimental Marine Biology and Ecology, 243(1): 115-135.
DOI:10.1016/S0022-0981(99)00113-6 |
Sufyan N, Mahamood M, Singh G K, et al. 2014. Description of Diplolaimelloides rushikondai sp. n. (Monhysteridae: Nematoda) and first report of the males of Myolaimus indicus (Myolaimidae: Nematoda) from India. International Journal of Nematology, 24(2): 141-148.
|
Timm R W. 1966. Some observations on the nematode genera Diplolaimella and Diplolaimelloides. Proceedings of Pakistan Academy of Science, 3(2): 114-125.
|
Tsalolikhin S Y. 1985. Nematodes of Fresh and Brackish Waters of Mongolia. Nauka Publishers, Leningrad. 115p. (in Russian)
|
Warwick R M, Platt H M, Somerfield P J. 1998. Free-living marine nematodes Part Ⅲ, Monhysterids. In: Barnes R S K, Crothers J H eds. Synopses of the British Fauna. Field Studies Council, London, UK. p. 14-22.
|
Warwick R M. 1981. The influence of temperature and salinity on energy partitioning in the marine nematode Diplolaimelloides bruciei. Oecologia, 51(3): 318-325.
DOI:10.1007/BF00540900 |