 2022, Vol. 40
2022, Vol. 40Institute of Oceanology, Chinese Academy of Sciences
Article Information
- ZHANG Yi, NI Mengqi, ZHENG Jinbin, CUI Zhaoxia
- Transcriptome of Eriocheir sinensis under air exposure
- Journal of Oceanology and Limnology, 40(2): 766-774
- http://dx.doi.org/10.1007/s00343-021-0449-7
Article History
- Received Nov. 19, 2020
- accepted in principle Jan. 25, 2021
- accepted for publication May. 12, 2021




2 Laboratory for Marine Biology and Biotechnology, Pilot Qingdao National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266071, China
The Chinese mitten crab, Eriocheir sinensis, is one of the most important commercial crustaceans in China. E. sinensis has the capability of being transported live without water, and the aquaculture industry of E. sinensis in China has gone through rapid development in the recent decade. In recent years, the annual aquaculture production of E. sinensis reached approximately 780 000 tons (MOA, 2020). Although the breeding industry of Chinese mitten crab has made great progress, the whole industrial chain of Chinese mitten crab still faces obstacles, especially during the transportation of Chinese mitten crab, and desiccation is an inevitable process. During this process, the ability of E. sinensis to deal with air exposure stress ensures long-distance transportation survival.
Air exposure is a common stress for aquatic animals, especially during their harvest and transportation (Ivanina et al., 2010). Thus, basic research on the air exposure stress response of the Chinese mitten crab has received growing attention. Furthermore, E. sinensis exhibits a relatively stronger tolerance to desiccation (Wang et al., 2013), which makes it a suitable species for investigating the molecular mechanism of air exposure stress response in aquatic crustaceans. Previous studies showed that E. sinensis could adjust their energy utilization mode, respiratory metabolism and antioxidant enzymes activities to adapt to air exposure stress (Bao et al., 2019a). Moreover, transcriptome analysis of gill provides insight into the immune and metabolism response of E. sinensis against air exposure stress (Bao et al., 2019b; Chen et al., 2019). However, the common and distinct mechanisms adopted by male and female crabs to cope with air exposure stress remain unclear.
With the development of high-throughput sequencing technologies, comparative transcriptome analysis has become a powerful method for investigating the molecular mechanisms of organisms under certain physiological status (Mutz et al., 2013). Recently, transcriptome analyses of gill have been performed to investigate the gene expression profiles of E. sinensis in response to air exposure (Bao et al., 2019b; Chen et al., 2019). However, different sexes of crabs could respond differently to environmental stress (Vitorino et al., 2019), thus, we performed a comparative transcriptome analysis the hepatopancreas rather than gill of female and male Chinese mitten crabs in response to air exposure stress, focusing on the transcriptional changes of metabolism, apoptosis, antioxidant enzymes, and heat shock proteins related genes. This study provides novel insights into the molecular mechanisms underlying the air exposure stress response in different sexes of E. sinensis.
2 MATERIAL AND METHOD 2.1 Air exposure and sample collectionCrabs (99.87±13.01 g in weight) were purchased from Dongying, Shandong Province. All crabs were acclimated for seven days in filtered aerated freshwater at a temperature of 22 ℃ and pH of 7.0. Crabs were fed once daily with clam until 24 h before beginning the experimental treatment. Preliminary experiments showed that crabs began to die at 30 h under air exposure, therefore, female/male crabs subjected to air exposure for 30 h were set as experimental groups (TR30FH/TR30MH) and crabs cultured in aerated freshwater served as control groups (TC30FH/ TC30MH). For both the control and air exposure groups, the experiment was run under the acclimation conditions of 22 ℃. The experiment was conducted in rectangular tanks (50 cm×35 cm×20 cm) and each group contained five individuals. After 30 h, the hepatopancreases of three individuals in each group of males and females were collected and were immediately frozen at -80 ℃ until the extraction of total RNA for analysis. No mortality was observed during the cultivation and analysis process.
2.2 Library construction and sequencingTotal RNA was extracted from hepatopancreas using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. The quantity and purity of total RNA were detected using NanoDrop ND-1000 (NanoDrop, Wilmington, DE, USA). The RNA integrity was assessed by Bioanalyzer 2100 (Agilent, CA, USA) with RIN number >7.0. Poly (A) RNA is purified from 1-μg total RNA using Dynabeads Oligo (dT)25-61005 (Thermo Fisher, CA, USA) using two rounds of purification. Then the poly(A) RNA was fragmented into small pieces using Magnesium RNA Fragmentation Module (NEB, cat.e6150, USA) under 94 ℃ for 5–7 min. Then the cleaved RNA fragments were reverse-transcribed to create the cDNA libraries using TruSeq Stranded mRNA Library Prep Kit (Illumina, San Diego, USA). Libraries were sequenced on an Illumina Hiseq 4000 (LC-Bio Technology Co., Ltd., Hangzhou, China) platform and 2×150 bp paired-end reads were generated.
2.3 Quality control and reads mappingRaw data were firstly subjected to quality control before bioinformatics analysis. In this step, reads containing sequencing adaptor, reads containing sequencing primer, and low-quality reads were removed. The sequence quality was verified using FastQC (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) including the Q20, Q30, and GCcontent of the clean data. All downstream analyses were based on clean data of high quality. The screened high-quality reads were mapped to the E. sinensis genome (Cui et al., 2021) using HISAT package (Kim et al., 2015). Then, all transcriptomes from samples were merged to reconstruct a comprehensive transcriptome using Perl scripts.
2.4 Identification of differentially expressed genesAfter the final transcriptome was generated, StringTie and Ballgown were used to estimate the expression levels of all transcripts. StringTie was used to perform expression level for mRNAs by calculating fragments per kilobase of transcript per million mapped reads (FPKM). Differential expression analysis was performed using the R package-Ballgown. Balldown was used for gene expression analysis (Frazee et al., 2015). Transcripts with |log2(fold change)|≥1 and P-values < 0.05 were considered DEGs.
2.5 Functional annotationFor functional annotation, all transcripts were searched against databases including the NCBI nonredundant protein sequences (NR) (http://www.ncbi.nlm.nih.gov/), Gene Ontology (GO) (http://www.geneontology.org/), and Kyoto Encyclopedia of Genes and Genomes (KEGG) (http://www.genome.jp/kegg/) databases using BlastX to convert nucleotide sequences into protein sequences (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
2.6 Validation of differentially expressed genes by quantitative real-time PCRSeveral differentially expressed genes, including phosphoenolpyruvate-carboxykinase (PEPCK), hexokinase, glutamine synthetase (GS), heat shock protein 70 (HSP70), HSP90, caspase, B-cell lymphoma-2 (Bcl2), and baculoviral IAP repeatcontaining protein 7 (BIRC7), were selected for validation by quantitative real-time PCR (qRT-PCR). Total RNA of hepatopancreas was extracted using RNAiso plus (TaKaRa, Japan), and reverse transcription was conducted using PrimeScriptTM RT reagent Kit with gDNA Eraser (TaKaRa, Japan) following the manufacturer's instructions. QRT-PCR was performed on an Applied Biosystems 7500 Realtime PCR System (Applied Biosystems, USA) using TB Green® Premix DimerEraserTM (TaKaRa, Japan) according to the manufacturer's instructions. Five biological and three technical replicates were performed, and β-actin served as the reference for internal standardization according to previous study (Yu et al., 2013). The specific primers for qRT-PCR were designed based on the sequences obtained from the transcriptome sequencing using Primer 5.0 software and are listed in Table 1. The reaction was conducted under the following conditions: 95 ℃ for 2 min, followed by 40 cycles of 95 ℃ for 15 s, 55 ℃ for 30 s and 72 ℃ for 30 s. Melting curve analysis was performed to evaluate the specificity of amplification. The 2-ΔΔCt method was used to calculate the relative mRNA expression levels as previously described (Livak and Schmittgen, 2001). Statistical significance between groups was analyzed using a one-way analysis of variance (ANOVA) followed by multiple comparison testing with the LSD-t test using the SPSS 17.0 software.

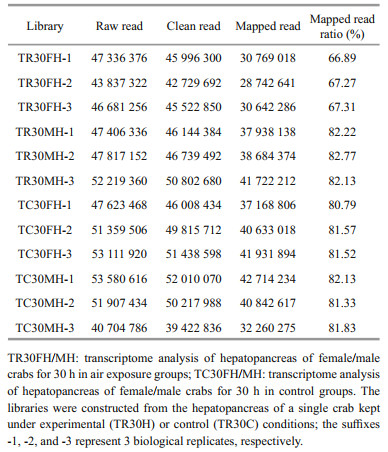
Based on the Illumina Hiseq 4000 sequencing platform, between 40 704 786–53 580 616 raw reads were generated from the 12 libraries (Table 2). After quality control, between 39 422 836–52 010 070 clean reads were obtained. The screened clean reads were mapped to the E. sinensis genome, and 28 742 641–42 714 234 mapped reads were identified. The transcriptome sequencing data has been submitted to the SRA database and the accession number is PRJNA699917.
In order to identify genes potentially involved in the air exposure stress responses of Chinese mitten crabs, the relative gene expression abundance was compared to identify DEGs of different treatments. A total of 2 186 and 4 813 DEGs were obtained in the hepatopancreas of female and male crabs under air exposure, respectively (Table 3). In detail, 956 up-regulated genes and 1 230 down-regulated genes were identified in female crabs exposed to air. Meanwhile, 433 up-regulated and 4 380 down-regulated genes were observed in male crabs under air exposure (Table 3).

To obtain comprehensive information on functions of DEGs, the transcripts were subjected to annotation analysis by searching against databases including NR, GO, and KEGG. In total, 428 and 1 322 transcripts were annotated. GO enrichment analysis showed that many DEGs identified in the TR30FH vs. TC30FH were enriched in categories of metabolism such as glycolytic process, glutathione metabolic process, and glutamine biosynthetic process (Fig. 1a). Meanwhile, several DEGs observed in the TR30MH vs. TC30MH were enriched in arginine biosynthetic process and citrate metabolic process (Fig. 1b). KEGG enrichment analysis showed that many DEGs identified in the TR30FH vs. TC30FH were enriched in metabolism processes including arachidonic acid metabolism, fatty acid biosynthesis, cholesterol metabolism, and steroid biosynthesis (Fig. 2a). Some DEGs detected in the TR30MH vs. TC30MH were enriched in metabolism pathways such as pyrimidine metabolism, cholesterol metabolism, nicotinate and nicotinamide metabolism, and steroid hormone biosynthesis (Fig. 2b). In the air exposure group of male and female crabs, several commonly expressed genes, associated with glycolysis, such as PEPCK, hexokinase, and lactate dehydrogenase (LDH), and associated with apoptosis, such as caspase, Bcl2 and BIRC7, showed the same expression trend (Fig. 3). Interestingly, glutathione peroxidase (Gpx) and catalase (CAT) were up-regulated in the air exposure group of male crabs, while up-regulation of HSP60, HSP70, and HSP90 occurred in air exposure female crabs (Fig. 3).

|
| Fig.1 Scatter diagram of Gene ontology (GO) enrichment for differentially expressed genes (DEGs) in E. sinensis a. TR30FH vs. TC30FH; b. TR30MH vs. TC30MH. Top 20 categories are listed in the scatter diagram. The magnitude of the dots displays gene number and the P-value is described by the color classification. The rich factor is a ratio of DEGs relative to all annotated genes considered in the GO. |

|
| Fig.2 Scatter diagram of KEGG pathway enrichment for DEGs a. TR30FH vs. TC30FH; b. TR30MH vs. TC30MH. Top 20 categories are listed in the scatter diagram. The magnitude of the dots displays gene number, and the P-value is described by the color classification. The rich factor is a ratio of DEGs relative to all annotated genes considered in the pathway. |

|
| Fig.3 Proposed scenario involving the air exposure response of E. sinensis ♀: female crabs; ♂: male crabs; ↑: gene expression level up-regulated; ↓: gene expression level down-regulated. |
To validate our sequencing results, several genes of interest were selected from the differentially expressed genes and verified by qRT-PCR. The results showed that PEPCK, GS, Hexokinase, Bcl2, BIRC7, HSP70, and HSP90 were up-regulated in the air exposure group of female crabs; PEPCK, GS, Hexokinase, Bcl2, and BIRC7 were up-regulated, and caspase were down-regulated in the air exposure group of male crabs. The differential expression tendencies of these genes detected by RNA-Seq and qRT-PCR is consistent (Fig. 4), indicating that the transcriptome data were reliable.

|
| Fig.4 Comparison of the relative fold changes of genes detected by RNA-seq and qRT-PCR in the hepatopancreas of female (a) and male (b) Chinese mitten crab Bars representing the RNA-seq data with * are values that were significantly different at P<0.05 from those of the control group. Bars representing the qRT-PCR data with * or ** indicate values (mean±SD) that were significantly different at P<0.05 or P<0.01 (n=5, ANOVA) from those of the control group. |
In crustaceans, the hepatopancreas plays essential roles in digestion, metabolism, detoxification, immune, and antioxidant defense, and has served as a crucial target tissue for assessing effects of environmental stress on organisms (Steenvoorden and van Henegouwen, 1997; Li et al., 2013; Cheng et al., 2020a). Air exposure led to a progressive degradation of the hepatopancreas tissue in Norway lobsters and caused the inflation of the liver tubules, and disappearance of connective tissue in black tiger shrimp (Ridgway et al., 2006; Duan et al., 2016). However, no information on the transcriptome analysis of hepatopancreas in E. sinensis in response to air exposure stress is available. Herein, a comparative transcriptome analysis of hepatopancreas was conducted to elucidate the common and distinct molecular mechanisms adopted by male and female E. sinensis to deal with air exposure stress.
Based on transcriptome sequencing and bioinformatics analysis, a total of 2 186 and 4 813 genes in the hepatopancreas of female and male crabs changed their expression levels in response to air exposure, implying that male crabs might be more sensitive to air exposure. To understand the possible functions of the DEGs, genes were searched against databases including NR, GO, and KEGG. However, approximately only less than one-third of the genes were annotated (Table 3), which may be related to the limited information available on the genetic background of crustaceans and arthropods (Li et al., 2012). Functional annotation showed that various genes such as heat shock proteins, antioxidant enzymes, apoptosis and glycolysis related genes participated in the air exposure response of E. sinensis.
Several common molecular mechanisms were found to be adopted by both male and female crabs to deal with air exposure stress (Fig. 3). Previous studies have revealed that air exposure stress could enhance glycolysis, which is the primary process of carbohydrate metabolism under acute hypoxic stress (Li et al., 2018; Sun et al., 2018). In this study, the transcriptional level of several key enzymes involved in glycolysis, including hexokinase, PEPCK, and LDH were significantly increased in the hepatopancreas of male and female E. sinensis under air exposure stress (Fig. 3). The up-regulated expression level of these enzymes implies an enhancement of glycolysis in the hepatopancreas of E. sinensis. Under the air exposure state, the crabs are unable to obtain enough oxygen, causing severe hypoxic stress and hindering aerobic respiration (Bao et al., 2019a). Consequently, anaerobic respiratory metabolism might be enhanced to provide sufficient energy for maintaining basic physiological activities. According to research, dietary carotenoids supplementation also may be a useful way for E. sinensis against air exposure (Liu et al., 2020).
Apoptosis is a normal biological process during the lifespan of organisms, and properly regulated apoptosis under stress is essential for maintaining homeostasis (Shan et al., 2021). Apoptotic pathways involve a set of proteins, such as caspases, tumor necrosis factor (TNF), p53 protein, Bcl2 family proteins and inhibitor of apoptosis proteins (IAPs) (Xie et al., 2020). In this study, the expression of caspase, a protein that promotes apoptosis, was decreased in the hepatopancreas of male and female E. sinensis subjected to air exposure stress (Fig. 3). Meanwhile, the transcriptional levels of two antiapoptotic proteins, i.e. Baculoviral IAP repeatcontaining protein 7 (BIRC7) and Bcl2 were increased in the hepatopancreas of E. sinensis regardless of gender (Fig. 3). The transcriptional alterations of these genes implied an inhibition of apoptosis in the hepatopancreas of E. sinensis suffering from air exposure stress. Our findings suggested that both male and female E. sinensis may inhibit cell apoptosis to resist air exposure stress.
Interestingly, distinct molecular responses in different sexes of E. sinensis were also observed. The expression levels of HSP60, HSP70, and HSP90 were increased in the hepatopancreas of female E. sinensis (Fig. 3), which was consistent with previous studies (Frenkel et al., 2008; Zhang et al., 2009; Huang et al., 2013; Lou et al., 2014). No significant transcriptional changes of these genes were observed in male E. sinensis. In this study, the expression levels of two important antioxidant enzymes, CAT and Gpx, were up-regulated in the hepatopancreas of male E. sinensis (Fig. 3), which was similar with findings in previous studies (Bao et al., 2019b; Cheng et al., 2020a). Similarly, no significant transcriptional changes of the two genes were observed in female E. sinensis.
HSPs, the well-known anti-stress proteins, generally act as molecular chaperones to regulate protein homeostasis by assisting protein folding, degrading misfolded protein, and preventing proteins from aggregation (Picard, 2002; Pratt and Toft, 2003; Kondrikov et al., 2015). HSPs have been reported to play essential roles in helping organisms to cope with diverse stress (John, 2009).
It has been reported that air exposure stress could result in overproduction of reactive oxygen species (ROS) and leads to oxidative damage (Sun et al., 2020; Cheng et al., 2020b). To prevent the toxicity caused by ROS and maintain proper cellular functions, organisms have developed effective antioxidant mechanisms to eliminate excessive ROS and maintain a balance of ROS (Jacob, 1995; Limón-Pacheco and Gonsebatt, 2009). Antioxidant enzymes are critical components of the antioxidant system (Davis and Pennypacker, 2017; Trivedi and Lal, 2017).
The up-regulations of HSPs and antioxidant enzymes are two crucial adaptive processes by which organisms deal with environmental stress (Abeyrathne et al., 2018; Bolhassani and Agi, 2019). Our study indicated that female E. sinensis might preferentially increase the expression of HSPs to cope with air exposure stress, while male E. sinensis tend to rely more on the up-regulation of antioxidant enzymes expression under air exposure stress.
5 CONCLUSIONBased on comparative transcriptome analysis, several putative molecular mechanisms adopted by different sexes of E. sinensis to cope with air exposure stress were preliminarily identified. Both female and male E. sinensis can make adaptative responses to air exposure via increasing the expression of glycolysis and anti-apoptotic related genes. Furthermore, female and male E. sinensis also up-regulated the transcriptional levels of HSPs and antioxidant enzymes, respectively, to cope with air exposure stress. This study enriches our knowledge on the molecular mechanisms underlying the air exposure stress response of E. sinensis.
6 DATA AVAILABILITY STATEMENTThe datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENTThe authors are grateful to all the laboratory members.
Abeyrathne E D N S, Huang X, Ahn D U. 2018. Antioxidant, angiotensin-converting enzyme inhibitory activity and other functional properties of egg white proteins and their derived peptides-a review. Poultry Science, 97(4): 1462-1468.
DOI:10.3382/ps/pex399 |
Bao J, Li X D, Xing Y N, Feng C C, Jiang H B. 2019a. Respiratory metabolism and antioxidant response in Chinese mitten crab Eriocheir sinensis during air exposure and subsequent reimmersion. Frontiers in Physiology, 10: 907.
DOI:10.3389/fphys.2019.00907 |
Bao J, Xing Y N, Jiang H B, Li X D. 2019b. Identification of immune-related genes in gills of Chinese mitten crabs(Eriocheir sinensis) during adaptation to air exposure stress. Fish & Shellfish Immunology, 84: 885-893.
|
Bolhassani A, Agi E. 2019. Heat shock proteins in infection. Clinica Chimica Acta, 498: 90-100.
DOI:10.1016/j.cca.2019.08.015 |
Chen X W, Wang J, Hou X, Yue W C, Li Z H, Wang C H. 2019. Gene expression profiles of gill provide insights into the aerial respiration capacity of the Chinese mitten crab, Eriocheir sinensis. Aquaculture, 506: 148-153.
DOI:10.1016/j.aquaculture.2019.03.027 |
Cheng C H, Ma H L, Deng Y Q, Feng J, Chen X L, Guo Z X. 2020a. Transcriptome analysis and histopathology of the mud crab (Scylla paramamosain) after air exposure. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 228: 108652.
|
Cheng J H, Lv X Y, Pan Y Y, Sun D W. 2020b. Foodborne bacterial stress responses to exogenous reactive oxygen species (ROS) induced by cold plasma treatments. Trends in Food Science & Technology, 103: 239-247.
|
Cui Z X, Liu Y, Yuan J B, Zhang X J, Ventura T, Ma K Y, Sun S, Song C W, Zhan D L, Yang Y, Liu H R, Fan G Y, Cai Q L, Du J, Qin J, Shi C C, Hao S J, Fitzgibbon Q P, Smith G G, Xiang J H, Chan T Y, Hui M, Bao C C, Li F H, Chu K H. 2021. The Chinese mitten crab genome provides insights into adaptive plasticity and developmental regulation. Nature Communications, 12: 2395.
DOI:10.1038/s41467-021-22604-3 |
Davis S M, Pennypacker K R. 2017. Targeting antioxidant enzyme expression as a therapeutic strategy for ischemic stroke. Neurochemistry International, 107: 23-32.
DOI:10.1016/j.neuint.2016.12.007 |
Duan Y F, Zhang Y, Dong H B, Zhang J S. 2016. Effect of desiccation on oxidative stress and antioxidant response of the black tiger shrimp Penaeus monodon. Fish & Shellfish Immunology, 58: 10-17.
|
Frazee A C, Pertea G, Jaffe A E, Langmead B, Salzberg S L, Leek J T. 2015. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nature Biotechnology, 33: 243-246.
DOI:10.1038/nbt.3172 |
Frenkel L, Dimant B, Portiansky E L, Maldonado H, Delorenzi A. 2008. Both heat shock and water deprivation trigger Hsp70 expression in the olfactory lobe of the crab Chasmagnathus granulatus. Neuroscience Letters, 443(3): 251-256.
DOI:10.1016/j.neulet.2008.07.072 |
Huang A M, Geng Y, Wang K Y, Zeng F, Liu Q, Wang Y, Sun Y, Liu X X, Zhou Y. 2013. Molecular cloning and expression analysis of heat shock protein 90 (Hsp90) of the mud crab, Scylla paramamosain. Journal of Agricultural Science, 5(7): 1-11.
|
Ivanina A V, Sokolov E P, Sokolova I M. 2010. Effects of cadmium on anaerobic energy metabolism and mRNA expression during air exposure and recovery of an intertidal mollusk Crassostrea virginica. Aquatic Toxicology, 99(3): 330-342.
DOI:10.1016/j.aquatox.2010.05.013 |
Jacob R A. 1995. The integrated antioxidant system. Nutrition Research, 15(5): 755-766.
DOI:10.1016/0271-5317(95)00041-G |
John T. 2009. Hsps and aging. Trends in Endocrinology & Metabolism Tem, 20(5): 216-222.
|
Kim D, Langmead B, Salzberg S L. 2015. HISAT: a fast spliced aligner with low memory requirements. Nature Methods, 12: 357-360.
DOI:10.1038/nmeth.3317 |
Kondrikov D, Fulton D, Dong Z, Su Y C. 2015. Heat shock protein 70 prevents hyperoxia-induced disruption of lung endothelial barrier via caspase-dependent and AIFdependent pathways. PloS One, 10: e0129343.
DOI:10.1371/journal.pone.0129343 |
Li C Z, Weng S P, Chen Y G, Yu X Q, Lü L, Zhang H Q, He J G, Xu X P. 2012. Analysis of Litopenaeus vannamei transcriptome using the next-generation DNA sequencing technique. PLoS One, 7: e47442.
DOI:10.1371/journal.pone.0047442 |
Li M X, Wang X D, Qi C L, Li E C, Du Z Y, Qin J G, Chen L Q. 2018. Metabolic response of Nile tilapia (Oreochromis niloticus) to acute and chronic hypoxia stress. Aquaculture, 495: 187-195.
DOI:10.1016/j.aquaculture.2018.05.031 |
Li X H, Cui Z X, Liu Y, Song C W, Shi G H. 2013. Transcriptome analysis and discovery of genes involved in immune pathways from hepatopancreas of microbial challenged mitten crab Eriocheir sinensis. PLoS One, 8: e68233.
DOI:10.1371/journal.pone.0068233 |
Limón-Pacheco J, Gonsebatt M E. 2009. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutation Research, 674(1-2): 137-147.
DOI:10.1016/j.mrgentox.2008.09.015 |
Liu M M, Jiang X D, Chen A Q, Chen T, Cheng Y X, Wu X G. 2020. Transcriptome analysis reveals the potential mechanism of dietary carotenoids improving antioxidative capability and immunity of juvenile Chinese mitten crabs Eriocheir sinensis. Fish & Shellfish Immunology, 104: 359-373.
|
Livak K J, Schmittgen T D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 25(4): 402-408.
DOI:10.1006/meth.2001.1262 |
Lou Y P, Li C, Landis A G, Wang G L, Stoeckel J, Peatman E. 2014. Transcriptomic profiling of differential responses to drought in two freshwater mussel species, the giant floater Pyganodon grandis and the Pondhorn Uniomerus tetralasmus. PLoS One, 9: e89481.
DOI:10.1371/journal.pone.0089481 |
MOA (Ministry of Agriculture). 2020. China Fishery Statistical Yearbook: 2020. China Agriculture Press, Beijing, China.
(in Chinese with English abstract)
|
Mutz K O, Heilkenbrinker A, Lönne M, Walter J G, Stahl F. 2013. Transcriptome analysis using next-generation sequencing. Current Opinion in Biotechnology, 24(1): 22-30.
DOI:10.1016/j.copbio.2012.09.004 |
Picard D. 2002. Heat-shock protein 90, a chaperone for folding and regulation. Cellular and Molecular Life Sciences, 59(10): 1640-1648.
DOI:10.1007/PL00012491 |
Pratt W B, Toft D O. 2003. Regulation of signaling protein function and trafficking by the hsp90/hsp70-based chaperone machinery. Experimental Biology and Medicine, 228(2): 111-133.
DOI:10.1177/153537020322800201 |
Ridgway I D, Taylor A C, Atkinson R J A, Stentiford G D, Chang E S, Chang S A, Neil D M. 2006. Morbidity and mortality in Norway lobsters, Nephrops norvegicus: physiological, immunological and pathological effects of aerial exposure. Journal of Experimental Marine Biology and Ecology, 328(2): 251-264.
DOI:10.1016/j.jembe.2005.07.015 |
Shan R T, Liu N, Yan Y Y, Liu B. 2021. Apoptosis, autophagy and atherosclerosis: relationships and the role of Hsp27. Pharmacological Research, 166: 105169.
DOI:10.1016/j.phrs.2020.105169 |
Steenvoorden D P T, van Henegouwen G M J B. 1997. The use of endogenous antioxidants to improve photoprotection. Journal of Photochemistry and Photobiology B: Biology, 41(1-2): 1-10.
DOI:10.1016/S1011-1344(97)00081-X |
Sun S M, Guo Z B, Fu H T, Zhu J, Ge X P. 2018. Integrated metabolomic and transcriptomic analysis of brain energy metabolism in the male Oriental river prawn(Macrobrachium nipponense) in response to hypoxia and reoxygenation. Environmental Pollution, 243: 1154-1165.
DOI:10.1016/j.envpol.2018.09.072 |
Sun Y, Lu Y F, Saredy J, Wang X W, Iv C D, Shao Y, Saaoud F, Xu K M, Liu M, Yang W Y, Jiang X H, Wang H, Yang X F. 2020. ROS systems are a new integrated network for sensing homeostasis and alarming stresses in organelle metabolic processes. Redox Biology, 37: 101696.
DOI:10.1016/j.redox.2020.101696 |
Trivedi S, Lal N. 2017. Antioxidant enzymes in periodontitis. Journal of Oral Biology & Craniofacial Research, 7(1): 54-57.
|
Vitorino H A, Alta R Y P, Ortega P. 2019. Lipid peroxidation in hepatopancreas, gill, and hemolymph of male and female crabs Platyxanthus orbignyi after air exposure. Journal of Marine Science and Engineering, 7(10): 347.
DOI:10.3390/jmse7100347 |
Wang M, Wang L, Zhou Z, Gao Y, Wang L, Shi X, Gai Y, Mu C, Song L. 2013. The molecular characterization of a catalase from Chinese mitten crab Eriocheir sinensis. International Journal of Immunogenetics, 40(3): 230-240.
DOI:10.1111/iji.12019 |
Xie Q Q, Liu Y, Li X F. 2020. The interaction mechanism between autophagy and apoptosis in colon cancer. Translational Oncology, 13(12): 100871.
DOI:10.1016/j.tranon.2020.100871 |
Yu A Q, Jin X K, Guo X N, Li S, Wu M H, Li W W, Wang Q. 2013. Two novel Toll genes (EsToll1 and EsToll2) from Eriocheir sinensis are differentially induced by lipopolysaccharide, peptidoglycan and zymosan. Fish & Shellfish Immunology, 35(4): 1282-1292.
|
Zhang X Y, Zhang M Z, Zheng C J, Liu J, Hu H J. 2009. Identification of two hsp90 genes from the marine crab, Portunus trituberculatus and their specific expression profiles under different environmental conditions. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, 150(4): 465-473.
|